






Por qué CB1 y CB2 importan en la ciencia del cannabis
GPCR Receptor acoplado a proteína G: un receptor de membrana que cambia de forma tras la unión de un ligando y transmite señales a través de socios intracelulares como las proteínas G y las beta-arrestinas.
Qué determina la salida del receptor
- Localización El tejido, el tipo celular y la posición subcelular moldean la respuesta.
- Identidad del ligando Los endocannabinoides, fitocannabinoides y ligandos sintéticos no impulsan estados de señalización idénticos.
- Socios disponibles Las distintas células ofrecen diferentes proteínas G, quinasas y beta-arrestinas.
- Patrón de exposición La duración de la señal y la estimulación repetida afectan la desensibilización y la internalización.
- Sesgo de vía Un ligando puede favorecer la señalización por proteína G, el reclutamiento de beta-arrestina u otras salidas.
Durante mucho tiempo, la explicación pública ha reducido la biología de los cannabinoid a una división ordenada: CB1 explica el “subidón”, CB2 se ocupa de la inflamación en algún lugar fuera del cerebro. Ese marco es demasiado tosco para ser útil. CB1 y CB2 son receptores acoplados a proteínas G, o GPCRs, y, como otros GPCRs, no actúan como simples interruptores de encendido y apagado. Traducen señales de endocannabinoid producidos por el organismo, phytocannabinoids de Cannabis sativa y ligandos sintéticos diseñados en laboratorios en respuestas celulares cambiantes. La respuesta que aparece depende de dónde se sitúe el receptor, qué ligando se una, qué socios de señalización estén disponibles, durante cuánto tiempo se estimule el receptor y si éste se orienta hacia la señalización por proteína G, el reclutamiento de β-arrestina, la desensibilización o la internalización.
Esto importa porque la ciencia del cannabis no trata solo de la intoxicación. También trata del dolor crónico, la epilepsia, la señalización inmunitaria, la neurodegeneración, el riesgo psiquiátrico y por qué tantos programas de fármacos basados en cannabinoid han parecido prometedores en la etapa preclínica y luego han tropezado en humanos. Hay mucho en juego. La Organización Mundial de la Salud estimó que 200 millones de personas usaron cannabis en 2019, alrededor del 4% de la población mundial de 15 a 64 años. La epilepsia afecta a unos 50 millones de personas en todo el mundo. La esquizofrenia afecta a unos 24 millones. Y, aun así, a fecha de 2025, la FDA de EE. UU. señala la aprobación de un producto farmacéutico derivado de cannabis y de tres productos farmacéuticos relacionados con cannabis. Esa brecha entre una exposición masiva y unos tratamientos aprobados limitados es una de las razones por las que la biología de los receptores importa tanto.
| Característica | CB1 | CB2 |
|---|---|---|
| Abreviatura típica | «receptor cerebral» | «receptor inmunitario» |
| Corrección del artículo | Enriquecimiento central, pero también expresión periférica | Enriquecido en el sistema inmunitario, pero no irrelevante para el cerebro |
| Funciones de ejemplo mencionadas | Percepción, memoria, control motor, nocicepción | Señalización de citocinas, migración celular, funciones neuroinflamatorias |
| Interpretación | Dependiente del circuito y del estado | Dependiente del tipo celular y del estado de la enfermedad |
Por qué la biología de los receptores explica más que las etiquetas de la planta
Las etiquetas como “indica”, “sativa” o incluso “dominante en THC” y “dominante en CBD” solo cuentan una parte de la historia, porque son los receptores, y no las categorías de mercadeo de la planta, los que están más cerca del mecanismo. El Δ9-tetrahydrocannabinol (THC) es un agonista parcial de CB1 y CB2, pero el efecto descendente de THC no es fijo. En neuronas corticales ricas en CB1, puede suprimir la liberación de neurotransmisores y alterar la percepción, la memoria y el control motor. En vías sensitivas periféricas, la misma familia de receptores puede modular la nocicepción. En células inmunitarias, la activación de CB2 puede modificar la señalización de citocinas o la migración celular. Misma familia. Distintos resultados.
La regla simple de que CB1 es solo cerebral y CB2 es solo inmunitario es demasiado burda para la biología actual de los receptores.Strong evidence
La vieja regla general — CB1 en el cerebro, CB2 en las células inmunitarias — partía de un patrón real, pero envejeció mal. La distribución es gradual y específica del tipo celular, no binaria. CB1 se expresa intensamente en muchas regiones del sistema nervioso central, especialmente en terminales presinápticas, aunque también aparece en tejidos periféricos. CB2 se asocia fuertemente con la función inmunitaria, pero ya no es defendible afirmar que es irrelevante para el cerebro. Una revisión de 2026 en Frontiers in Behavioral Neuroscience sostuvo que la señalización de CB2 ha cobrado atención en trastornos del sistema nervioso central, en concreto a través de mecanismos neuroinflamatorios y neurodegenerativos, y describió esto como “una actualización de los últimos 3 años”. Esa actualización importa. Si CB2 contribuye a la patología central en algunas condiciones, entonces los fármacos dirigidos a CB2 no pueden entenderse como herramientas puramente periféricas.
La estructura profundiza la historia. Una revisión de 2026 en Frontiers in Chemical Biology explicó que la selectividad de los ligandos entre “CB1 y CB2” surge de diferencias estructurales a nivel del receptor que influyen en la pose de unión, la eficacia y la regulación del receptor. En términos simples, pequeños cambios químicos pueden sesgar un ligando hacia un subtipo de receptor o hacia una vía de señalización concreta, lo que ayuda a explicar por qué dos cannabinoid que parecen similares sobre el papel pueden sentirse o actuar de forma muy distinta in vivo. Un estudio indexado en PubMed de 2025/2026 sobre selectividad de subtipo fue más allá al mostrar que la selectividad de los endocannabinoid está vinculada a la dinámica conformacional del receptor y no a un modelo rígido de llave-cerradura. El receptor se mueve. El ligando estabiliza algunos estados más que otros. La biología sigue esos estados.
De los phytocannabinoids a la señalización endocannabinoide
{kind=link}
Señalización retrógrada Un patrón de señalización sináptica en el que una célula postsináptica libera un mensajero que viaja hacia atrás para actuar sobre receptores presinápticos.
Secuencia del descubrimiento en breve
- 1 Se aisló y se definió estructuralmente THC.
- 2 Se demostraron sitios específicos de unión a cannabinoides en tejido cerebral.
- 3 CB1 se clonó como GPCR.
- 4 CB2 se identificó a partir de tejidos relacionados con la inmunidad.
- 5 La anandamida y luego el 2-AG establecieron un sistema de señalización endógeno.
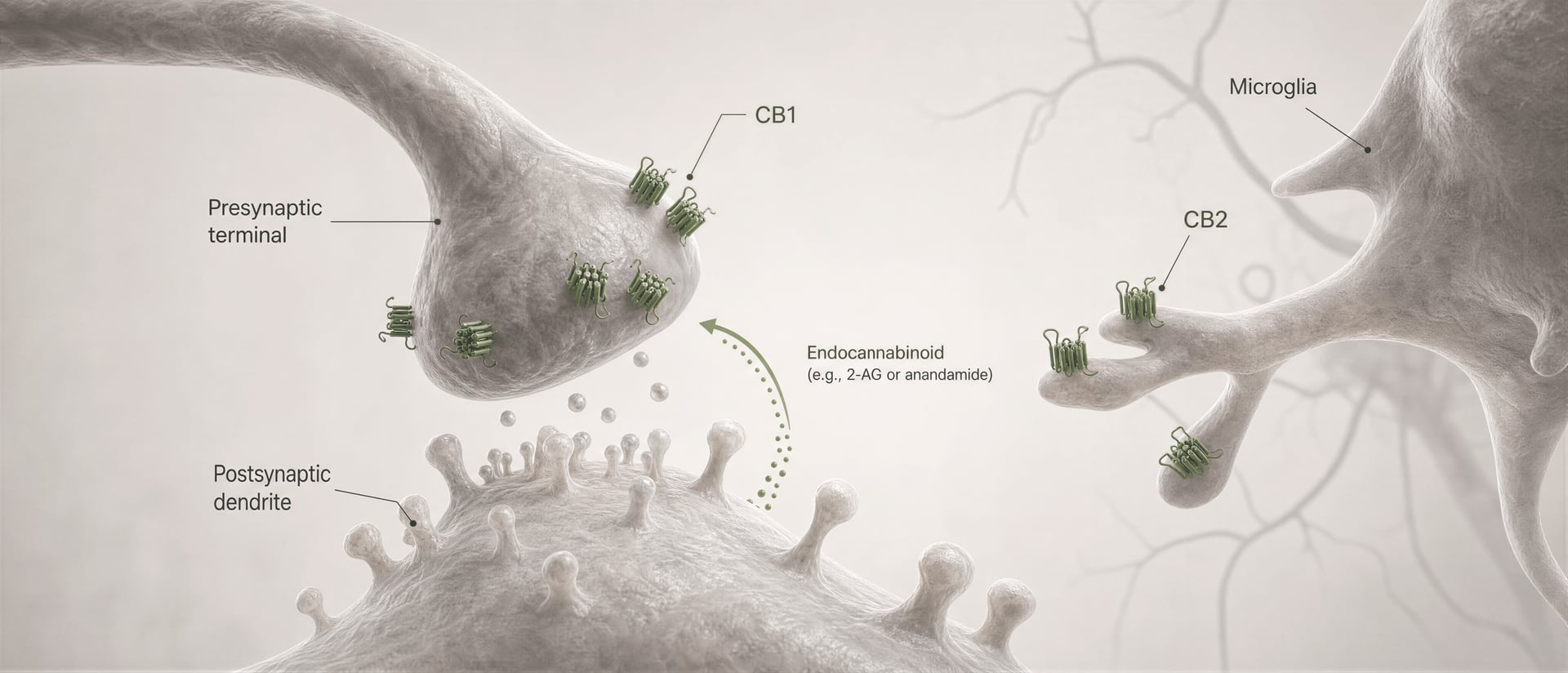
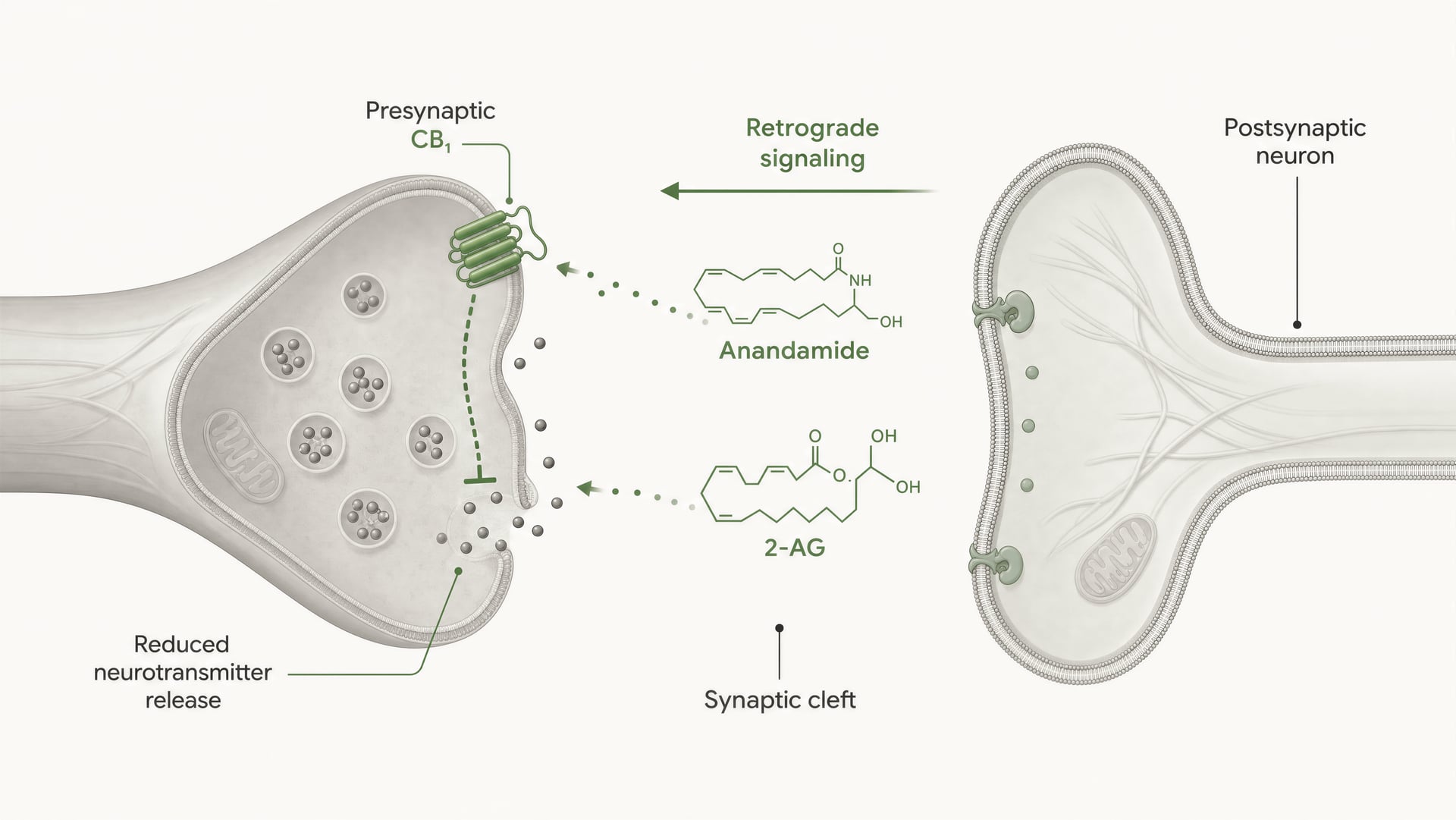
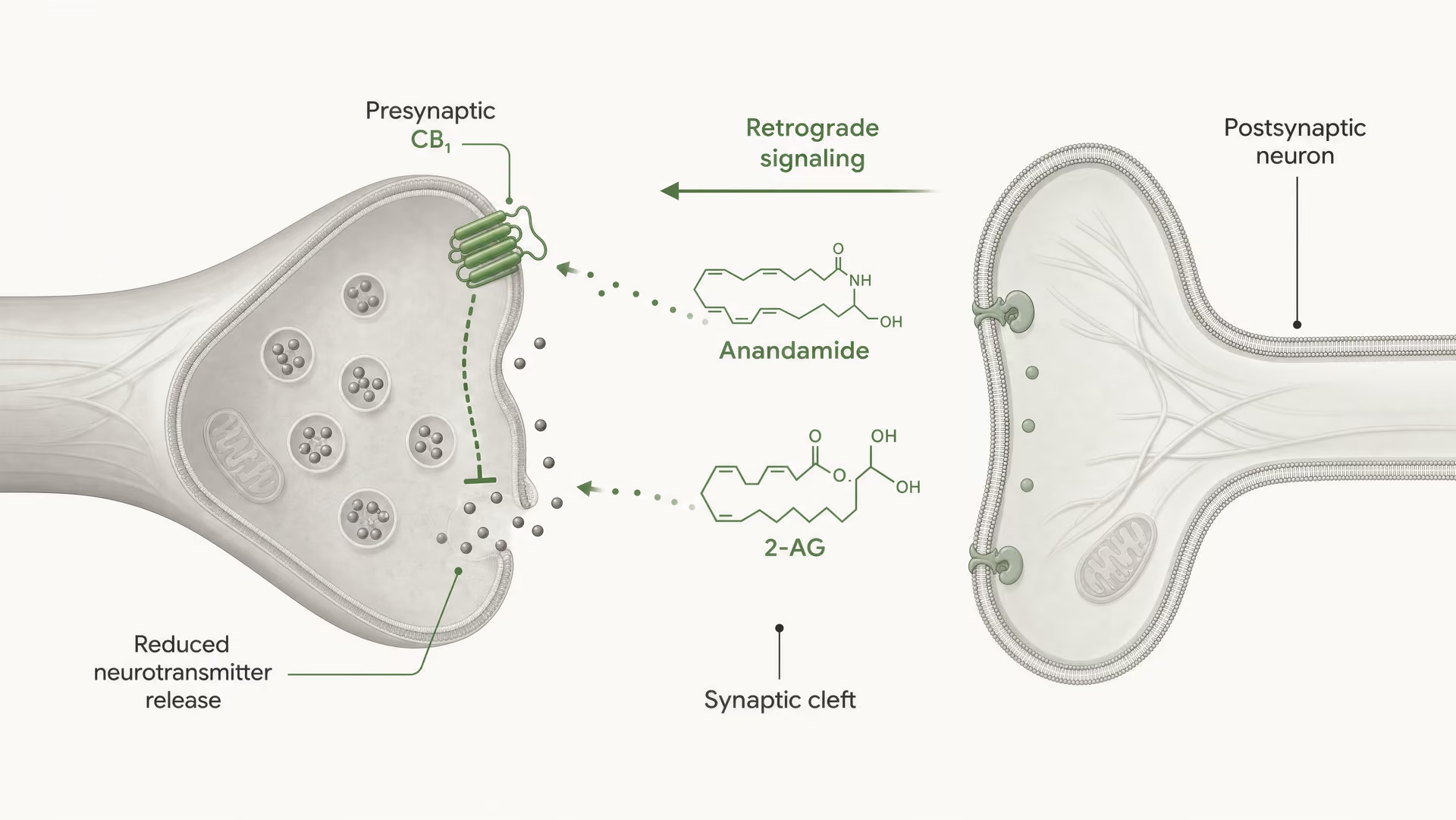
Los receptores cannabinoid no se descubrieron porque el organismo evolucionara para cannabis. La secuencia fue la contraria. El trabajo de Allyn Howlett y sus colegas fue fundamental para definir la farmacología de los receptores cannabinoid, y el descubrimiento posterior de la anandamida por Raphael Mechoulam y Lumír Hanuš ayudó a establecer que los seres humanos producen sus propias moléculas de señalización similares a cannabinoid. La anandamida y el 2-arachidonoylglycerol, normalmente abreviado como 2-AG, son los principales endocannabinoid. Se producen “a demanda”, no se almacenan en vesículas como los neurotransmisores clásicos y, con frecuencia, señalan de forma retrógrada: una célula postsináptica genera un endocannabinoid que viaja hacia atrás a través de la sinapsis para activar CB1 presináptico y reducir la liberación ulterior de neurotransmisores.
Eso representa una imagen fundamentalmente distinta de “una sustancia de la marihuana golpea un receptor”. La señalización endocannabinoide es local, transitoria y está estrechamente regulada por enzimas de síntesis y degradación. Los phytocannabinoids entran en ese sistema desde fuera. Los ligandos sintéticos pueden afectarlo con mayor intensidad o de manera más selectiva. El resultado es que el mismo receptor puede ser activado por un pulso endógeno efímero, por un phytocannabinoid oral de absorción lenta o por un agonista sintético de alta eficacia con responsabilidades de seguridad muy distintas.
Esta diferencia es una de las razones por las que la intoxicación no puede inferirse solo a partir del nombre del receptor. Depende de la eficacia del ligando, la dosis, la vía, el momento y el contexto tisular. THC en CB1 es central para los efectos psicoactivos, sí, pero ese hecho no reduce a CB1 a un “receptor de la psicoactividad”. Tampoco convierte a CB2 en un simple regulador antiinflamatorio. El artículo de 2025 en American Journal of Psychiatry sobre señalización sesgada de CB1 hizo precisamente este punto más amplio al argumentar que los ligandos sesgados hacia CB1 podrían ofrecer una estrategia terapéutica para la esquizofrenia. Esa propuesta vincula la ciencia del cannabinoid con una idea más amplia de GPCR: si un ligando favorece ramas de señalización beneficiosas y evita otras vinculadas a efectos adversos, la acción del fármaco podría separarse de la activación burda del receptor. Si esa promesa se mantiene clínicamente sigue sin resolverse, pero el argumento mecanístico es sólido.
Qué significa este artículo por distribución, señalización y dianas farmacológicas
En este artículo, distribución significa algo más que un mapa de órganos. Incluye densidad del receptor, tipo celular, localización subcelular, estado de la enfermedad y cambio temporal. Un receptor expresado en terminales GABAérgicas puede tener efectos de circuito distintos de los del mismo receptor en terminales glutamatérgicas. Un receptor regulado al alza durante la inflamación no es equivalente a su estado basal. La distribución es dinámica.
Señalización significa las consecuencias intracelulares del compromiso del receptor. Para CB1 y CB2, esto incluye el acoplamiento a proteínas G de la familia Gi/o, la inhibición de la adenilil ciclasa, la modulación de canales iónicos, cambios en cascadas de quinasas, el reclutamiento de β-arrestina, la desensibilización del receptor y la internalización. También incluye la modulación alostérica y el agonismo sesgado, en los que los ligandos pueden favorecer algunas salidas de señalización sobre otras. Esto no es una nota al pie académica. Con frecuencia, es la diferencia entre analgesia, sedación, tolerancia, disforia o un ensayo fallido.
Dianas farmacológicas significa receptores considerados para intervención, no historias de éxito garantizadas. La focalización selectiva de CB1 podría reducir algunos efectos fuera de diana, pero aun así topar con eventos adversos centrales. La focalización selectiva de CB2 puede evitar algunas responsabilidades intoxicantes, pero la selectividad no garantiza eficacia en la compleja enfermedad humana. El trabajo de biología de sistemas deja eso claro. Un análisis integrador de redes indexado en PubMed de 2025/2026 identificó CB1 y CB2 como nodos altamente influyentes en el sistema endocannabinoid y relacionó su señalización con vías metabólicas más amplias. En otras palabras, estos receptores forman parte de redes mayores. Si se empuja un nodo, otras vías se desplazan.
Esa es la postura de este artículo. CB1 y CB2 son nodos de señalización dependientes del contexto. No son interruptores estáticos. No son meras etiquetas para “cerebro” y “sistema inmunitario”. Si la ciencia del cannabis va a explicar por qué un compuesto parece terapéutico en un contexto, intoxicante en otro y decepcionante en la clínica, debe comenzar en el nivel del receptor y permanecer allí el tiempo suficiente para seguir la biología hasta donde realmente va.
Una breve historia del descubrimiento del receptor cannabinoide
Antes de que se identificaran los receptores cannabinoides, la ciencia del cannabis era sobre todo una historia de química. Los investigadores podían aislar compuestos de la planta, comparar efectos conductuales crudos en animales y discutir la potencia, pero todavía no podían explicar cómo una molécula como delta-9-tetrahidrocannabinol, o THC, producía sus efectos con una precisión comparable a la de un receptor. Eso cambió a finales de la década de 1980 y principios de la de 1990. El cambio fue decisivo: la investigación sobre cannabis pasó de catalogar fitocannabinoides a estudiar interacciones ligando-receptor, señalización intracelular, distribución tisular y, finalmente, el sistema lipídico endógeno que hoy se denomina sistema endocannabinoide, o ECS.
| Año | Hito | Personas mencionadas en el artículo |
|---|---|---|
| 1964 | Aislamiento y estructura de THC | Raphael Mechoulam; Yechiel Gaoni |
| 1988 | Sitios de unión a cannabinoides específicos de alta afinidad en membranas cerebrales de rata | Allyn Howlett; William Devane |
| 1990 | Clonación de CB1 | Lisa Matsuda y colegas |
| 1992 | Identificación de la anandamida | William Devane; Lumír Hanuš; Raphael Mechoulam; colegas |
| 1993 | Identificación de CB2 | Sean Munro; Kerrie Thomas; M. Abu-Shaar |
| 1995 | Identificación de 2-AG por grupos separados | Equipo de Mechoulam; grupo de Sugiura |
De la farmacología del THC a la identificación del receptor
Un hito temprano clave se produjo en 1964, cuando Raphael Mechoulam y Yechiel Gaoni informaron del aislamiento y la estructura del THC. Ese logro fue importante porque dio a los farmacólogos una molécula definida para probar, en lugar de un extracto botánico variable. Durante las dos décadas siguientes, el campo construyó un mapa de relación estructura-actividad a partir del THC y cannabinoides relacionados, pero seguía existiendo debate sobre el mecanismo. Algunos investigadores favorecían efectos inespecíficos sobre la membrana porque los cannabinoides son lipofílicos. Esa postura se volvió más difícil de defender a medida que se acumulaban datos de unión estereoselectiva y saturable.
La era de los receptores comenzó propiamente con estudios de unión en la década de 1980. En 1988, Allyn Howlett y William Devane publicaron en Molecular Pharmacology un artículo histórico que mostraba sitios de unión cannabinoide específicos y de alta afinidad en membranas cerebrales de rata utilizando el agonista sintético CP55,940. El resultado no fue una vaga sugerencia de una diana. Mostraba saturabilidad, variación regional y especificidad farmacológica compatibles con un receptor auténtico. El tejido cerebral no respondía a los cannabinoides como si estos simplemente se disolvieran en las bicapas lipídicas y alteraran todo a la vez. Había selectividad.
Tres años después, en 1990, Lisa Matsuda y colaboradores clonaron el primer receptor cannabinoide, ahora llamado CB1, y lo publicaron en Nature. CB1 fue identificado como un receptor acoplado a proteína G, o GPCR, un hallazgo que situó de inmediato la farmacología cannabinoide dentro de una de las superfamilias de señalización más importantes de la biología. Eso era relevante porque los GPCR no son solo interruptores. Adoptan múltiples estados conformacionales, se acoplan a distintos socios intracelulares, se desensibilizan, se internalizan y muestran sesgo de señalización dependiente del ligando. Esas ideas cobrarían centralidad mucho más tarde, pero la clonación de CB1 las hizo posibles.
CB2 siguió rápidamente. En 1993, Sean Munro, Kerrie Thomas y M. Abu-Shaar identificaron un segundo receptor cannabinoide, CB2, también en Nature, caracterizado inicialmente a partir de tejidos relacionados con el sistema inmunitario. Este descubrimiento creó una abreviatura duradera que dio forma al campo durante años: CB1 como el “receptor cerebral” asociado con la intoxicación, CB2 como el receptor “periférico” o inmunitario asociado con la inflamación. La abreviatura fue útil, pero ya entonces era demasiado grosera, y ha envejecido mal. La distribución de ambos receptores depende de la especie, el tipo celular, el estado de activación, el contexto de enfermedad y el método de ensayo.
Cómo CB1 y CB2 cambiaron el campo del endocannabinoid
Una vez identificados CB1 y CB2, la siguiente pregunta obvia fue por qué el cuerpo tenía receptores para cannabinoides de origen vegetal. La respuesta llegó en 1992, cuando William Devane, Lumír Hanuš, Raphael Mechoulam y colaboradores identificaron la anandamida, formalmente araquidonoiletanolamida, como un ligando endógeno. El artículo, publicado en Science, marcó una ruptura conceptual. La farmacología del cannabis ya no trataba solo de compuestos exógenos de Cannabis sativa. Trataba de un sistema lipídico de señalización nativo.
Un segundo ligando endógeno importante, el 2-araquidonoilglicerol o 2-AG, fue identificado en 1995 por grupos independientes, incluidos el equipo de Mechoulam y el grupo de Sugiura. Con receptores y ligandos endógenos en su lugar, el ECS se expandió rápidamente. Los investigadores identificaron enzimas de síntesis y degradación como la amida hidrolasa de ácidos grasos, FAAH, para la anandamida, y la lipasa de monoacilgliceroles, MAGL, para el 2-AG. También se enfrentaron a una cuestión aún no resuelta: cómo estas moléculas altamente lipofílicas se desplazan a través de las membranas y del espacio extracelular. El campo suele hablar de “transporte”, pero un transportador endocannabinoide único y dedicado ha seguido siendo esquivo.
Este fue el punto en el que la ciencia cannabinoide dejó de ser un esquema de dos receptores y pasó a ser una red de señalización. CB1 y CB2 se vincularon a proteínas Gi/o, a la inhibición de la adenilil ciclasa, a la modulación de canales de calcio y potasio y a la supresión de la liberación de neurotransmisores. Pero la historia no se quedó ahí. Los receptores podían reclutar beta-arrestinas, sufrir desensibilización e internalización y responder de manera diferente a fitocannabinoides, endocannabinoid y ligandos sintéticos incluso cuando esos ligandos, en teoría, actuaban sobre el mismo receptor. El lenguaje actual de agonismo sesgado de los GPCR encaja especialmente bien con los cannabinoides. Un artículo de 2025 en American Journal of Psychiatry argumentó que la señalización sesgada de CB1 es una estrategia terapéutica plausible para la esquizofrenia, un trastorno que afecta a unos 24 millones de personas en todo el mundo según la WHO. Eso está muy lejos de la antigua imagen de CB1 como mero receptor que explica por qué el THC produce intoxicación.
La historia de CB2 también se ha ampliado. Los trabajos iniciales lo situaron sobre todo en tejidos inmunitarios, y eso era direccionalmente correcto, pero estudios posteriores hallaron expresión de CB2 en microglía y, bajo ciertas condiciones, en otras poblaciones celulares del sistema nervioso central. Una revisión de 2026 en Frontiers in Behavioral Neuroscience describió “an update over the last 3 years” vinculando la señalización de CB2 con mecanismos neuroinflamatorios y neurodegenerativos, dejando claro que CB2 no puede descartarse como irrelevante para el cerebro. El trabajo estructural actual ha ido aún más lejos. Una revisión de 2026 en Frontiers in Chemical Biology sobre “CB1 and CB2” destacó que la selectividad de subtipo depende de diferencias estructurales a nivel del receptor que alteran la unión, la eficacia y la regulación. Un estudio reciente indexado en PubMed sobre selectividad de subtipo sostiene asimismo que la selectividad de los endocannabinoid es dinámica, moldeada por el comportamiento conformacional más que por un modelo simple de llave y cerradura.
Investigadores fundamentales y por qué la historia sigue importando
Tres nombres ocupan un lugar central en esta historia. Raphael Mechoulam ayudó a definir la base química y biológica de la ciencia cannabinoide, desde el trabajo sobre la estructura del THC hasta el descubrimiento del endocannabinoid. Lumír Hanuš fue una figura central en la identificación de la anandamida y en la investigación posterior sobre endocannabinoid. La farmacología de receptores de Allyn Howlett fue decisiva para demostrar que los cannabinoides actúan a través de sitios de unión y mecanismos de señalización específicos en el cerebro. Sin su trabajo, no existiría el campo moderno del ECS.
La historia sigue importando porque las simplificaciones antiguas siguen distorsionando los debates actuales. En 2019, se estimó que 200 millones de personas en todo el mundo, aproximadamente el 4% de las personas de 15 a 64 años, consumían cannabis según la WHO. Al mismo tiempo, la FDA afirma que ha aprobado un producto farmacéutico derivado del cannabis y tres productos farmacéuticos relacionados con cannabis. La exposición pública es enorme. La traslación clínica es selectiva y difícil. La historia de los receptores explica por qué. Los efectos de cannabinoid dependen de la clase de ligando, del estado del receptor, de la localización tisular, del momento y del sesgo de la vía. También dependen de una red más amplia. Un análisis integrador de red de 2025/2026 identificó CB1 y CB2 como nodos altamente influyentes conectados con vías metabólicas más que como dianas aisladas.
Ese es el verdadero legado del descubrimiento de los receptores. No simplificó la biología del cannabis. Mostró por qué la biología es más compleja de lo que la antigua división entre cerebro y cuerpo permitía.
Dónde se encuentra CB1: circuitos cerebrales, tejidos periféricos y gradientes funcionales
CB1 se ganó su reputación como el principal receptor cannabinoide psicoactivo por una razón. Abunda en el sistema nervioso central, y el trabajo de farmacología de receptores de Allyn Howlett ayudó a establecer que el THC actúa a través de un sistema receptor específico y saturable, en lugar de mediante efectos inespecíficos sobre la membrana. Pero la antigua abreviatura —CB1 en el cerebro, CB2 en las células inmunitarias— ahora genera más confusión que claridad. CB1 está fuertemente enriquecido en circuitos neuronales, sí. Pero también está presente en el intestino, el hígado, el tejido adiposo, los órganos reproductores, los tejidos cardiovasculares y las vías sensoriales, donde moldea la ingesta, el metabolismo, la señalización del dolor y la función autónoma. La distribución es amplia. La función es condicional.
Eso importa porque la exposición a cannabinoid es común. La World Health Organization estimó que 200 millones de personas usaron cannabis en 2019, aproximadamente el 4% de la población mundial de 15 a 64 años. También importa porque la farmacología de receptores sigue entrando en medicina: la U.S. FDA afirma que tiene aprobado un producto farmacéutico derivado del cannabis y tres productos farmacéuticos relacionados con cannabis. Un receptor presente en tantos órganos no puede reducirse a una sola etiqueta conductual.
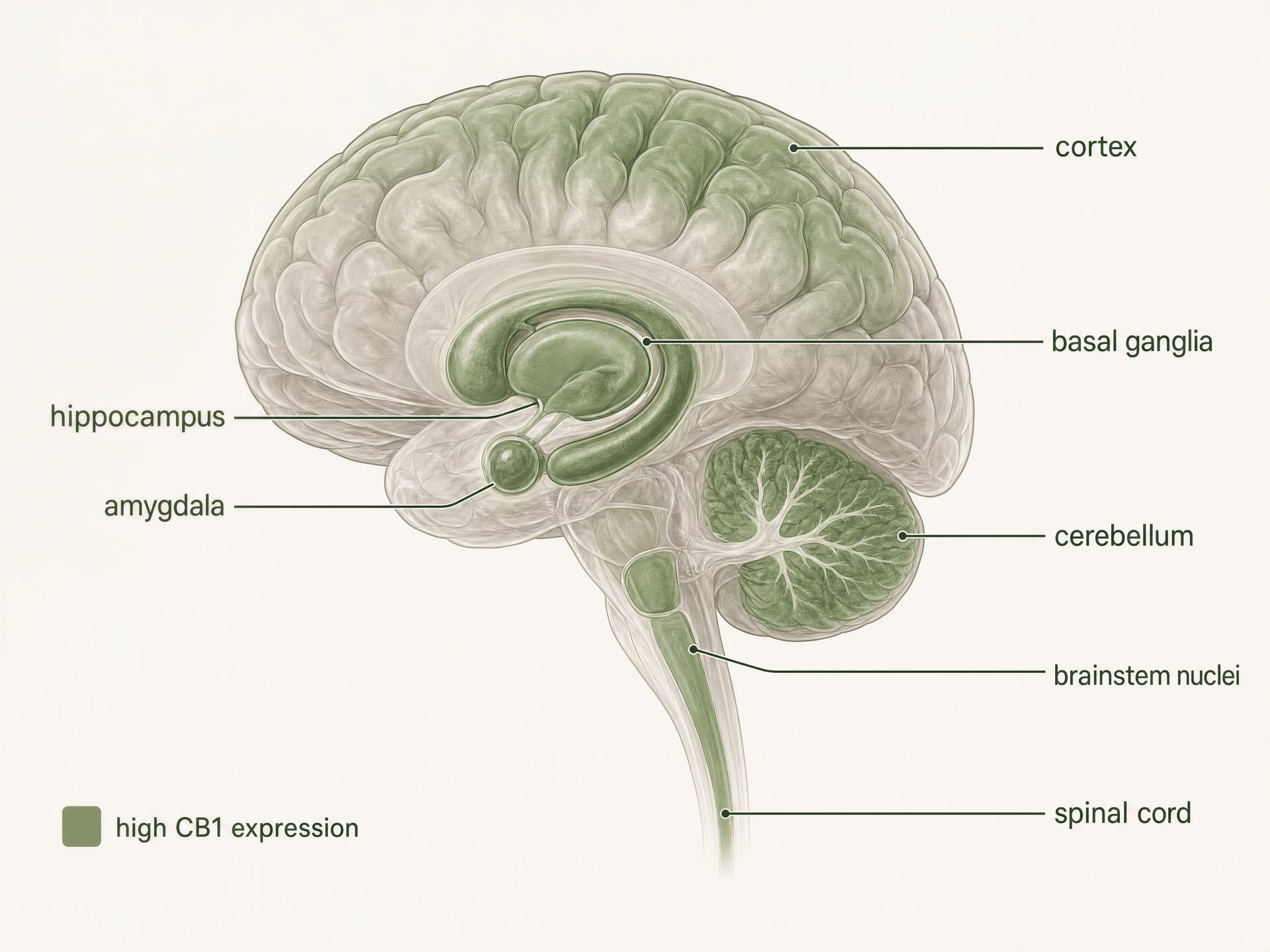
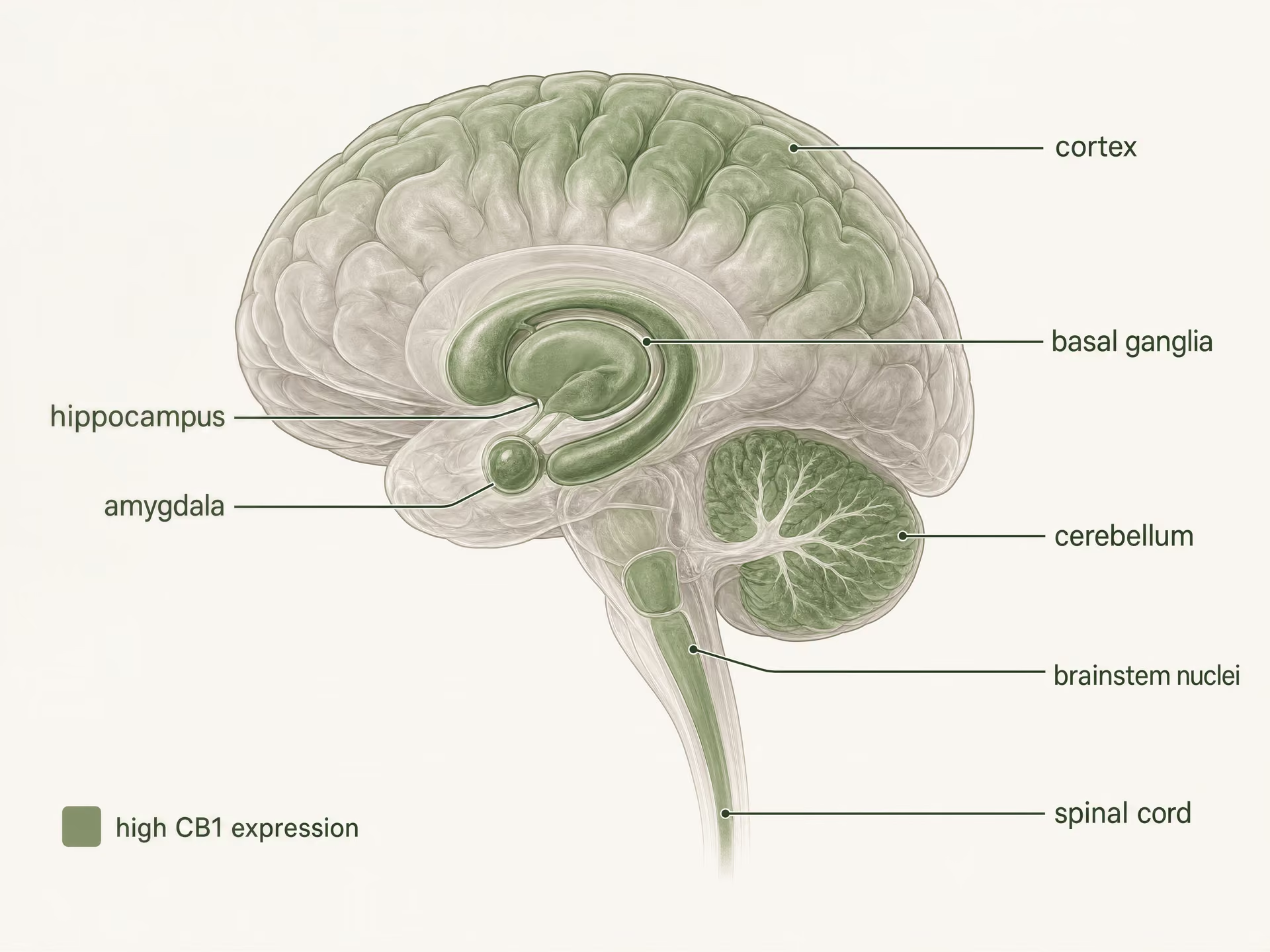
- Patrón general
- Uno de los GPCR más abundantes en el cerebro de los mamíferos
- Regiones de alta densidad nombradas
- Corteza, hipocampo, amígdala, ganglios basales, cerebelo
- Sitios relacionados con el dolor nombrados
- Sustancia gris periacueductal, médula ventromedial rostral, asta dorsal
- Sitios periféricos nombrados
- Intestino, hígado, tejido adiposo, vías reproductivas, cardiovasculares y sensoriales
Alta expresión en el sistema nervioso central
{kind=link}
CB1 es uno de los receptores acoplados a proteína G más abundantes en el cerebro de los mamíferos. La autorradiografía, la hibridación in situ y el mapeo inmunohistoquímico construyeron una imagen clara mucho antes de los estudios estructurales actuales: aparecen altas densidades en la corteza, el hipocampo, la amígdala, los ganglios basales, el cerebelo y varias regiones implicadas en el procesamiento del dolor, con expresión adicional en núcleos del tronco encefálico y a lo largo de la médula espinal. Este patrón encaja sorprendentemente bien con los efectos clásicos del THC, aunque no de forma perfecta.
| Localización de CB1 | Efecto sináptico inmediato | Consecuencia de ejemplo nombrada en el artículo |
|---|---|---|
| Terminales de interneuronas GABAérgicas | Suprime la liberación de GABA | Desinhibición de neuronas descendentes |
| Terminales glutamatérgicas | Suprime la liberación de glutamato | Disminución de la excitación |
| Circuitos de ganglios basales y cerebelo | Altera la liberación de neurotransmisores en vías motoras | Enlentecimiento motor, alteración de los circuitos de hábito, coordinación deteriorada |
| Vías del dolor | Modula la transmisión nociceptiva | Cambios en el procesamiento del dolor ascendente, descendente, inflamatorio y afectivo |
En la corteza y el hipocampo, CB1 se sitúa en circuitos que regulan la atención, la memoria de trabajo, el aprendizaje de extinción y la plasticidad sináptica a corto plazo. Los efectos sobre la memoria no son simplemente “hipocampo igual a olvido”. Dependen en gran medida de qué terminales axónicas expresan el receptor. CB1 suele concentrarse presinápticamente, donde suprime la liberación de neurotransmisores tras la activación por endocannabinoid como la anandamida y el 2-araquidonoilglicerol, los lípidos de señalización cuyo descubrimiento por Raphael Mechoulam, Lumír Hanuš y sus colegas transformó el campo. Cuando CB1 se activa en terminales de interneuronas GABAérgicas, puede desinhibir neuronas aguas abajo; cuando se activa en terminales glutamatérgicas, puede amortiguar la excitación. Mismo receptor, resultado de red opuesto.
Los ganglios basales y el cerebelo explican otro conjunto familiar de efectos. La expresión densa de CB1 en el estriado, el globo pálido, la sustancia negra pars reticulata y las capas moleculares del cerebelo vincula la activación del receptor con enlentecimiento motor, alteración de los circuitos del hábito, coordinación deteriorada y, en algunas dosis, efectos similares a la catalepsia en modelos animales. Sin embargo, el hecho de que CB1 sea escaso en los centros cardiorrespiratorios del tronco encefálico en comparación con los receptores opioides ayuda a explicar por qué la sobredosis de cannabinoid no suele producir el mismo patrón de depresión respiratoria fatal observado con agonistas opioides potentes. La ubicación importa. También importa lo que está ausente.[5]The Health Effects of Cannabis and Cannabinoids: The Current State of Evidence and Recommendations for Research. National Academies of Sciences, Engineering, and Medicine. National Academies Press, 2017. https://nap.nationalacademies.org/catalog/24625/the-health-effects-of-cannabis-and-cannabinoids-the-current-state
El procesamiento del dolor muestra la misma lógica regional. CB1 se encuentra en la sustancia gris periacueductal, la médula rostral ventromedial, el asta dorsal de la médula espinal y las vías nociceptivas periféricas. Eso le da al receptor múltiples puntos de entrada en la nocicepción: puede alterar las señales ascendentes del dolor, el control descendente del dolor, la sensibilización inflamatoria y la carga emocional del dolor. Esta es una de las razones por las que los cannabinoid han permanecido en la discusión sobre dolor crónico, especialmente desde que casi 1 de cada 5 adultos en los Estados Unidos vive con dolor crónico, según las National Academies. Pero la analgesia no está garantizada simplemente porque CB1 esté presente. La sedación, el deterioro cognitivo, la tolerancia y los efectos adversos limitantes por dosis suelen aparecer a través de circuitos cercanos o de los mismos circuitos en distintos niveles de ocupación del receptor.
La señalización sesgada de CB1 puede separar los efectos terapéuticos deseados de los efectos psicoactivos o cognitivos indeseados.Limited evidence
Proteínas Gi/o Una familia de proteínas G que comúnmente reducen la actividad de la adenilil ciclasa y ayudan a controlar los canales iónicos tras la activación del GPCR.
La biología moderna de los receptores añade otra capa. CB1 no es un simple interruptor de encendido y apagado. Se acopla principalmente a proteínas Gi/o, reduciendo la actividad de la adenilil ciclasa y modulando canales iónicos, pero también puede reclutar beta-arrestinas, sufrir desensibilización e internalización, y mostrar sesgo de señalización dependiente del ligando. El artículo de 2025 del American Journal of Psychiatry que sostiene que la señalización sesgada de CB1 podría explotarse terapéuticamente en la esquizofrenia lo dice de forma directa: la ocupación del receptor por sí sola es un mal predictor del resultado. Con la esquizofrenia afectando a unos 24 millones de personas en todo el mundo, el atractivo de separar la señalización deseada de los efectos psicoactivos o cognitivos no deseados es evidente. Si esa separación es alcanzable en la práctica sigue siendo una cuestión abierta de desarrollo de fármacos, no un hecho establecido.
CB1 periférico en intestino, hígado, tejido adiposo y más allá
CB1 fuera del cerebro no es una nota al pie. Es central para entender por qué los cannabinoid afectan el apetito, las náuseas, el manejo de la glucosa, el metabolismo lipídico y la sensibilidad visceral.
En el intestino, CB1 se expresa en el sistema nervioso entérico, en compartimentos epiteliales y en vías relacionadas con el vago. Su activación puede ralentizar el vaciamiento gástrico, alterar la motilidad intestinal, reducir el vómito y cambiar la señalización entre el intestino y el cerebro. Los efectos sobre el apetito a menudo se describen como si se originaran únicamente en los centros hipotalámicos de recompensa y alimentación, pero el CB1 periférico contribuye a la historia al moldear las entradas sensoriales y hormonales antes incluso de que las señales lleguen a esos circuitos. Una comida no actúa sobre un paisaje receptor en blanco; cambia el tono de endocannabinoid localmente.
En el hígado y el tejido adiposo, CB1 participa en la regulación metabólica, incluida la lipogénesis, la sensibilidad a la insulina y el almacenamiento de energía. Esta fue una de las grandes lecciones de la era de rimonabant. Bloquear CB1 mejoró el peso y los marcadores metabólicos, lo que respaldó la idea de que una señalización de endocannabinoid hiperactiva contribuye a la fisiopatología de la obesidad. Pero rimonabant, un inverso agonista de CB1 con actividad central, también produjo efectos adversos psiquiátricos graves, incluida depresión y ansiedad, y fue retirado. Ese episodio suele citarse como un fracaso del “direccionamiento hacia CB1”. Más exactamente, fue un fracaso de un tipo particular de direccionamiento hacia CB1: un antagonismo central intenso o inverso agonismo en un sistema receptor integrado en circuitos de estado de ánimo y estrés. La lección no es que el CB1 periférico sea irrelevante; es que el patrón de exposición al fármaco y el estado del receptor importan tanto como el nombre del receptor.
Adipocitos, hepatocitos, tejido pancreático, músculo esquelético, tejidos cardiovasculares y órganos reproductivos también amplían el mapa periférico. Lo mismo ocurre con las neuronas sensoriales. El análisis de red integrador indexado en PubMed de 2025/2026 que identificó CB1 y CB2 como nodos altamente influyentes en la señalización de endocannabinoid es útil aquí porque desplaza el marco desde la mera localización del receptor hacia su participación en redes metabólicas y de señalización. Un receptor con expresión modesta en un tejido aún puede ejercer efectos sistémicos grandes si ocupa un cuello de botella en la señalización local.
El trabajo estructural también mantiene honesta esta discusión. La revisión de 2026 en Frontiers in Chemical Biology sobre CB1 y CB2 enfatiza que la selectividad y la eficacia de los ligandos surgen de diferencias estructurales a nivel del receptor que alteran la unión, la señalización y la regulación del receptor. Un estudio indexado en PubMed de 2025/2026 sobre la selectividad de subtipo también sostiene que la dinámica conformacional, y no solo el ajuste llave-cerradura, da forma a cómo los endocannabinoid distinguen los subtipos de receptor. Eso importa para la distribución de CB1 porque “CB1 en el hígado” no significa que THC, anandamida, 2-AG y un agonista sintético hagan todos lo mismo allí.
Por qué la distribución no equivale a una función única y uniforme
El mayor error en los mapas de receptores es tratar la expresión como destino. No lo es. La alta expresión indica dónde mirar, no qué ocurrirá.
Primero, el tipo celular cambia el signo del efecto. Un receptor CB1 en una terminal glutamatérgica puede reducir la excitación. El mismo receptor en una terminal GABAérgica puede reducir la inhibición. Esos no son resultados intercambiables. Segundo, la localización sináptica importa. CB1 suele ser presináptico, a menudo activado por endocannabinoid liberados “a demanda” por neuronas postsinápticas, creando un control retrógrado sobre la liberación de neurotransmisores. Esa disposición favorece una modulación breve, dependiente de la actividad, más que una activación constante del receptor.
Tercero, la identidad del ligando importa. Los endocannabinoid son mensajeros locales de corta vida. Los fitocannabinoid como THC llegan desde fuera del sistema, a menudo con exposiciones más altas y más duraderas que las señales endógenas. Los ligandos sintéticos pueden empujar aún más, con distinta eficacia y sesgo. Algunos promueven con más fuerza la señalización Gi/o; otros favorecen el reclutamiento de beta-arrestinas, la desensibilización o la internalización del receptor. Por eso dos compuestos pueden llamarse agonistas de CB1 y, sin embargo, diferir marcadamente en la estimulación del apetito, la alteración de la memoria, el deterioro motor y el desarrollo de tolerancia.
Cuarto, la disponibilidad local del ligando lo cambia todo. La anandamida y el 2-AG se producen y degradan en el sitio, por lo que sus efectos dependen de la actividad neuronal, el estado metabólico, la expresión enzimática y el contexto inflamatorio. Quinto, la densidad del receptor en sí existe en un gradiente. La región cerebral, la etapa del desarrollo, el estado de enfermedad y la exposición repetida a fármacos modifican los niveles de CB1 y su respuesta.
La literatura actual se está alejando de las dicotomías precisamente por esta razón. La revisión de 2026 en Frontiers in Behavioral Neuroscience señala una actualización en los últimos 3 años sobre cómo se entiende la señalización de los receptores cannabinoid en los trastornos del SNC, especialmente cuando se incluyen mecanismos neuroinflamatorios y neurodegenerativos. CB1 debe leerse con la misma cautela. Es un receptor central dominante, pero no exclusivamente central; un receptor de la alimentación, pero no solo eso; una diana para el dolor, pero no un interruptor analgésico limpio. Cualquier descripción seria de la distribución de CB1 tiene que pensar en gradientes, circuitos y estados de señalización, en lugar de en una caricatura cerebro-versus-cuerpo.
Dónde se encuentra CB2: raíces del sistema inmunitario y el mapa creciente del SNC
La vieja fórmula lo decía con claridad: CB1 es el receptor cerebral, CB2 es el receptor inmunitario. Ese marco ayudó en la enseñanza inicial, pero ahora confunde más de lo que aclara. CB2 sí muestra un enriquecimiento clásico fuera de las neuronas, especialmente en linajes inmunitarios y hematopoyéticos, y ese hecho sigue importando para la farmacología. Sin embargo, la literatura más reciente, en especial la revisión de 2026 de Frontiers in Behavioral Neuroscience, plantea una afirmación más fuerte: CB2 ahora se discute en trastornos del sistema nervioso central porque su expresión y señalización se hacen más visibles en microglía, circuitos inflamatorios y estados relacionados con lesión, una “actualización de los últimos 3 años” que ha cambiado la forma de mapear e interpretar el receptor. El resultado no es que CB2 de repente se haya convertido en un receptor cerebral de alta abundancia y pan-neuronal. No lo hizo. El resultado es que la distribución del receptor debe describirse como condicional, específica de célula y dependiente del estado.
Esa distinción importa clínicamente. La World Health Organization estimó que 200 millones de personas usaron cannabis en 2019, es decir, el 4% de la población mundial de 15 a 64 años. Incluso con solo un pequeño número de medicamentos relacionados con cannabinoid aprobados —la FDA contabilizó en 2025 un producto derivado de cannabis y tres productos relacionados con cannabis—, la localización del receptor sigue moldeando dónde buscan los desarrolladores de fármacos efectos antiinflamatorios, analgésicos, neuroprotectores y psiquiátricos, y dónde esperan toxicidades.
- Enriquecimiento clásico
- Células inmunitarias y hematopoyéticas
- Tipos celulares nombrados
- Linfocitos B, linfocitos T, macrófagos, monocitos, células asesinas naturales, neutrófilos, mastocitos
- Tejidos canónicos nombrados
- Bazo, amígdala, timo, médula ósea, células inmunitarias circulantes
- Relevancia del SNC enfatizada
- Microglía y estados vinculados con patología
Enriquecimiento clásico en células inmunes y hematopoyéticas
CB2 se identificó originalmente como el subtipo de receptor cannabinoide con la expresión más fuerte en células vinculadas a la inmunidad más que a la transmisión sináptica rápida. Ese sigue siendo el punto de partida correcto. En comparación con CB1, que está ampliamente representado en muchas poblaciones neuronales, CB2 está clásicamente enriquecido en linfocitos B, linfocitos T, macrófagos, monocitos, células asesinas naturales, neutrófilos, mastocitos y otros compartimentos hematopoyéticos. Por ello, el bazo, las amígdalas, el timo, la médula ósea y las poblaciones circulantes de células inmunitarias han sido los tejidos canónicos para el análisis de CB2.
Este sesgo hacia el sistema inmunitario moldeó la idea temprana de desarrollo de fármacos de los agonistas selectivos de CB2 como una forma de captar beneficios antiinflamatorios o analgésicos mientras se evitaban los efectos intoxicantes asociados con una fuerte activación de CB1 en el cerebro. Fue una hipótesis sensata, pero solo parcialmente completa. CB2 es un GPCR acoplado a Gi/o, y al igual que CB1 no simplemente se “enciende” o se “apaga”. Según el ligando, la conformación del receptor y el contexto celular, CB2 puede reducir la actividad de la adenilil ciclasa, influir en las vías MAPK, alterar de forma indirecta el acoplamiento de canales iónicos, reclutar beta-arrestinas y sufrir desensibilización o internalización. Así que incluso en tejidos inmunes periféricos, la verdadera cuestión no es solo si CB2 está presente, sino qué células lo expresan, en qué nivel, bajo qué estímulo y con qué sesgo descendente.
Esa complejidad es una de las razones por las que ligandos de aspecto similar pueden comportarse de manera distinta. La revisión de 2026 en Frontiers in Chemical Biology sobre la estructura del receptor cannabinoide sostiene que la selectividad en “CB1 y CB2” está determinada por diferencias estructurales a nivel del receptor que alteran la unión del ligando, la eficacia de señalización y la regulación del receptor. Un estudio indexado en PubMed de 2025/2026 sobre selectividad de subtipo llevó la idea más lejos al mostrar que la selectividad del endocannabinoid es dinámica y está vinculada a estados conformacionales, no a un modelo rígido de llave-cerradura. Esto importa para el mapeo tisular porque un ligando endógeno como 2-AG o anandamida, un fitocannabinoid como THC y un agonista sintético con preferencia por CB2 pueden encontrarse con la misma población de receptores, pero estabilizar salidas de señalización diferentes.
Por tanto, el mapa inmunocéntrico más antiguo de CB2 no era erróneo. Era incompleto. CB2 sigue describiéndose mejor como un receptor con fuertes raíces en el sistema inmunitario. Pero las raíces no son el organismo completo.
| Contexto | Cómo se describe CB2 | Punto interpretativo |
|---|---|---|
| Base cerebral sana | A menudo bajo o cercano a los límites de detección en muchas regiones | Una señal basal baja no equivale a ausencia de relevancia |
| Microglía activada | Más detectable tras lesión o inflamación | Sostiene la relevancia del SNC mediante funciones de tipo inmunitario |
| Astrocitos / endoteliales / células infiltrantes | Se ha informado en algunos contextos de enfermedad | La localización depende del método y del modelo |
| Expresión neuronal constitutiva amplia | Necesita pruebas más sólidas | El artículo trata esta afirmación con cautela |
CB2 en microglía, neuroinflamación y estados de lesión
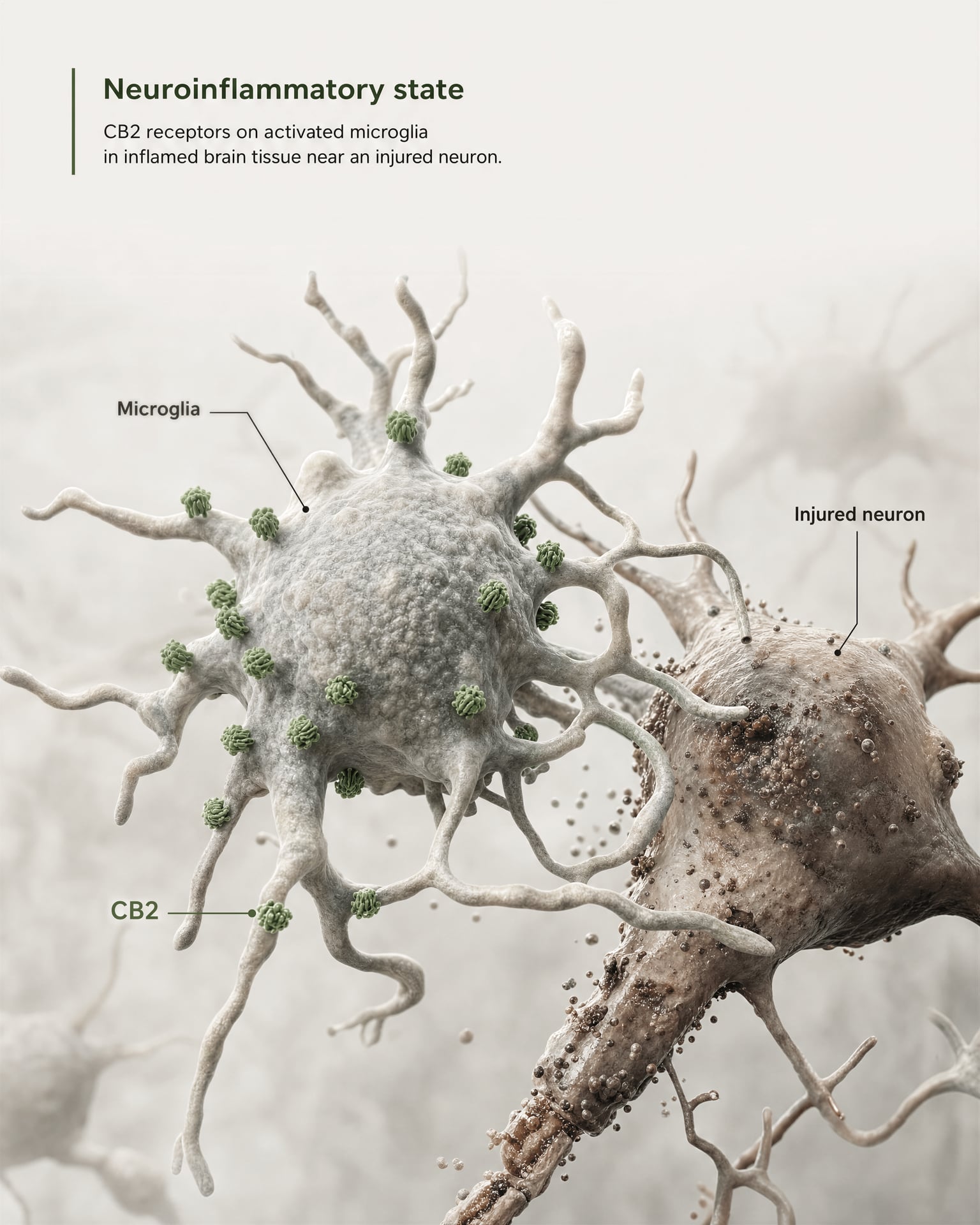
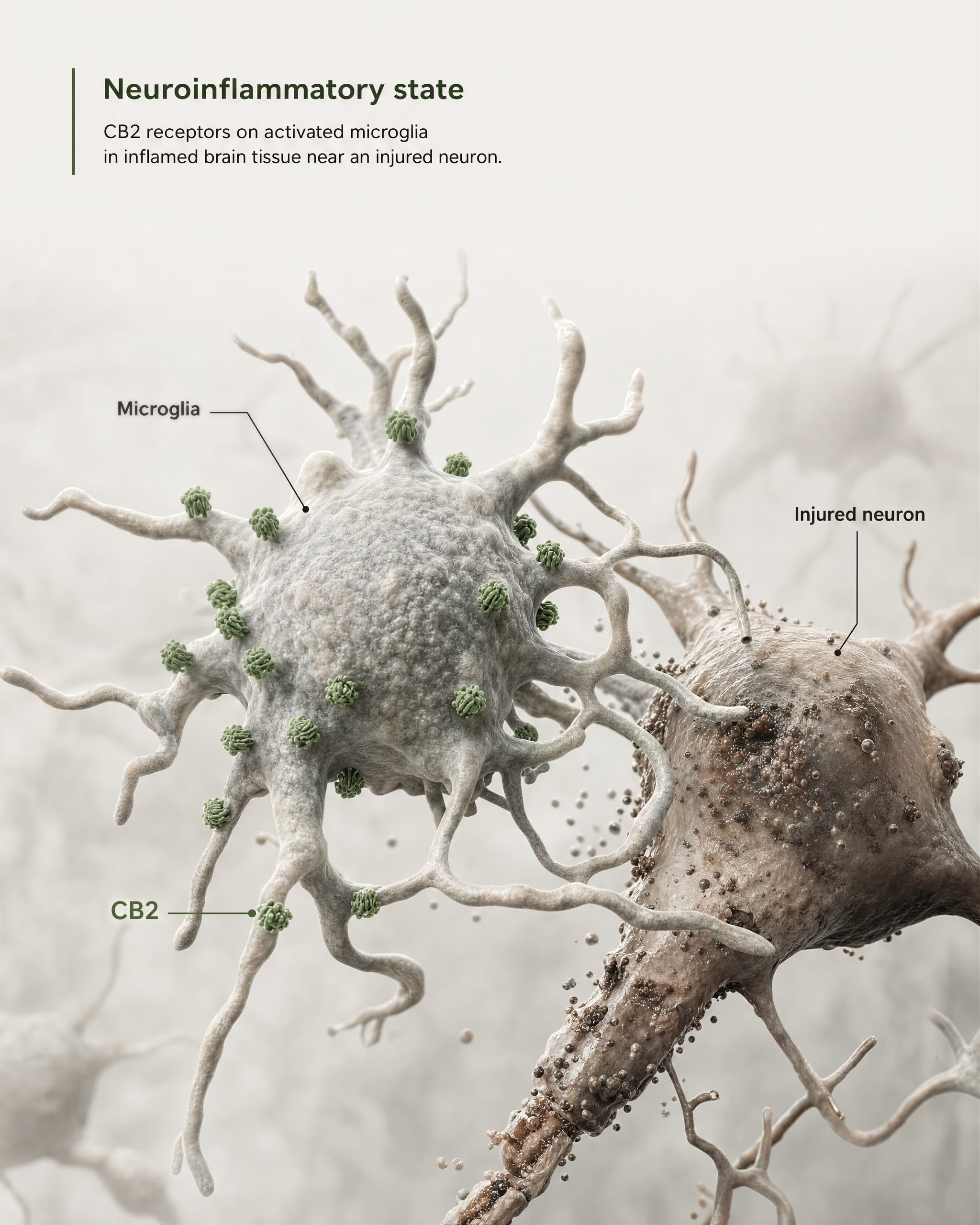
El argumento más sólido a favor de la relevancia central no proviene de afirmar que CB2 esté ampliamente abundante en neuronas sanas del prosencéfalo. Proviene de la microglía y de la biología de la enfermedad.
{kind=link}
La microglía son las células inmunitarias residentes del SNC, y se sitúan exactamente en la frontera donde el antiguo modelo de “receptor inmune periférico” empieza a fallar. Si un receptor se expresa en el sistema propio de vigilancia inmunitaria y respuesta inflamatoria del cerebro, entonces llamarlo meramente periférico deja de ser exacto. La revisión de 2026 en Frontiers in Behavioral Neuroscience plantea este punto de forma directa: la señalización de CB2 ha ganado atención en trastornos del SNC porque se vincula con mecanismos neuroinflamatorios y neurodegenerativos. Por eso CB2 aparece ahora en debates sobre la enfermedad de Alzheimer, la enfermedad de Parkinson, la esclerosis múltiple, la lesión cerebral traumática, el ictus, el dolor neuropático y algunas afecciones psiquiátricas en las que la señalización inflamatoria forma parte de la patología.
La frase clave es expresión inducida o aumentada. En muchas regiones cerebrales sanas, la expresión basal de CB2 es baja, a veces cercana a los límites de los métodos de detección más antiguos. Pero tras una lesión, una infección, una inflamación crónica o una neurodegeneración, la señal de CB2 suele volverse más detectable, en particular en microglía activada y, en algunos estudios, en astrocitos, células inmunitarias infiltrantes, compartimentos endoteliales o subconjuntos neuronales restringidos. Esta es una regla de distribución muy distinta de la que suele aplicarse a CB1. CB1 suele estar constitutivamente presente en abundancia en circuitos neuronales definidos. CB2 se interpreta con más frecuencia como un receptor cuya relevancia en el SNC emerge bajo estrés, patología o activación inflamatoria.
Esa distinción tiene consecuencias prácticas. Un fármaco dirigido a CB2 puede tener poco efecto en un tejido sano donde la densidad del receptor es baja, pero mostrar una actividad medible en un tejido enfermo donde la expresión ha aumentado y las redes de señalización han cambiado. Esta inducibilidad es una de las razones por las que los hallazgos preclínicos han sido a la vez prometedores y difíciles de trasladar. El momento importa. El estadio de la enfermedad importa. La composición celular importa. Un entorno microglial poslesión no es farmacológicamente equivalente a un corte cerebral no estimulado.
Los problemas interpretativos no son triviales. CB2 tiene una larga historia de preocupaciones sobre la especificidad de los anticuerpos, detección de transcritos de baja abundancia, diferencias entre especies y afirmaciones inconsistentes de localización entre métodos. Algunos informes tempranos probablemente sobrestimaron CB2 neuronal porque las herramientas disponibles eran débiles. Por eso, los estudios cuidadosos ahora se apoyan en evidencias convergentes —transcriptómica de célula única, hibridación in situ, reporteros genéticos validados, controles de knockout, datos proteómicos cuando es posible y comparaciones dependientes del estado— en lugar de un único resultado de tinción. Si un estudio informa de CB2 en neuronas en basal y otro no logra detectarlo, la discrepancia puede reflejar diferencias regionales reales, estado de enfermedad, especie, edad o, simplemente, limitaciones del ensayo.
CB2 tiene una relevancia significativa para el SNC, especialmente en contextos gliales e inflamatorios vinculados con lesión.Limited evidence
Por tanto, la mejor posición actual es contenida pero clara: CB2 tiene una relevancia real en el SNC, principalmente a través de funciones gliales e inmunoafines, y esa relevancia aumenta durante la neuroinflamación y la lesión. Las afirmaciones sobre una expresión neuronal constitutiva amplia de CB2 en el cerebro normal requieren pruebas más sólidas que las afirmaciones sobre CB2 microglial y asociado a la patología.
Cómo cambiaron las últimas 3 años la conversación sobre CB2
La revisión de 2026 en Frontiers in Behavioral Neuroscience enmarca explícitamente la literatura reciente como una “actualización de los últimos 3 años”, y esa formulación captura un cambio real. La conversación pasó de debatir si CB2 está “en el cerebro” a secas a preguntar dónde, cuándo y en qué estados de enfermedad su señalización se vuelve aprovechable.
Tres desarrollos impulsaron ese cambio. Primero, mejoraron los métodos de resolución celular. Los conjuntos de datos de RNA de célula única y de núcleo único, un mapeo espacial mejor y estándares de validación más estrictos redujeron la posibilidad de que la expresión de bajo nivel o inducible se descartara simplemente porque los ensayos antiguos carecían de sensibilidad. Segundo, la neuroinflamación pasó a ocupar un lugar central en muchos modelos de trastornos cerebrales. Una vez que las enfermedades se analizaron a través de mecanismos inmunitarios y gliales en lugar de marcos centrados solo en neuronas, CB2 se volvió mucho más difícil de ignorar. Tercero, la propia farmacología de receptores maduró. El campo piensa ahora más en términos de eficacia, sesgo de señalización, tráfico del receptor y respuestas dependientes del contexto que en una simple ocupación del receptor.
Esa perspectiva más amplia de GPCR es visible incluso fuera de la literatura sobre CB2. El artículo de 2025 en American Journal of Psychiatry sobre señalización sesgada de CB1 y esquizofrenia sostiene que la farmacología cannabinoide debe entenderse a través de la señalización sesgada y no de la activación cruda del receptor. La esquizofrenia afecta a unos 24 millones de personas en todo el mundo, según WHO, por lo que no se trata de una cuestión académica marginal. La misma lógica se aplica a CB2. Un ligando “selectivo de CB2” en el papel puede seguir produciendo resultados distintos según si impulsa preferentemente la señalización de proteínas G, el reclutamiento de beta-arrestina, la internalización del receptor o programas transcripcionales antiinflamatorios en microglía activada.
La visión sistémica más reciente refuerza esto. Un estudio de análisis de redes indexado en PubMed de 2025/2026 identificó a CB1 y CB2 como nodos altamente influyentes en el endocannabinoid system y conectó la señalización de los receptores con vías metabólicas en lugar de aislar los receptores del resto de la biología celular. Eso coincide con lo que muestran los datos de CB2 en el SNC: la distribución no es una entrada fija de atlas. Forma parte de una red de señalización adaptativa.
La conclusión es simple. CB2 debe seguir presentándose como un receptor cannabinoide enriquecido en el sistema inmunitario. Pero detenerse ahí ahora da una imagen incorrecta. En el cerebro, CB2 se entiende mejor como un receptor de baja expresión basal, inducible y vinculado a la enfermedad, cuya importancia se vuelve más clara en la microglía y en los estados neuroinflamatorios, y cuya detección sigue dependiendo en gran medida del método, el modelo y el momento.
Cómo señalan los receptores cannabinoides: acoplamiento Gi/o, segundos mensajeros y efectos sinápticos
La farmacología de los receptores cannabinoides comienza con una afirmación simple que se complica rápidamente: CB1 y CB2 son receptores acoplados a proteínas G de clase A, y ambos señalan con mayor frecuencia a través de proteínas Gi/o. Ese hecho básico, establecido mediante trabajos fundacionales sobre receptores realizados por Allyn Howlett y otros, sigue siendo válido. Lo que ha cambiado es la comprensión de lo que realmente significa el acoplamiento Gi/o en las células. No significa un único efecto descendente. Significa un conjunto de efectos posibles cuya combinación depende del ligando, la densidad del receptor, el estado de fosforilación, el entorno de la membrana, el tipo celular y el momento.
Esa distinción importa porque aproximadamente 200 millones de personas usaron cannabis en 2019, o el 4% de la población mundial de 15 a 64 años según la Organización Mundial de la Salud, mientras que la FDA afirma que, a fecha de 2025, hay un producto farmacológico derivado de cannabis y tres productos farmacológicos relacionados con cannabis aprobados. La señalización del receptor no es una cuestión secundaria. Es el mecanismo que separa un fármaco antiseizure útil de la sedación, un fármaco para el apetito que fracasa de los efectos adversos psiquiátricos, y un ligando selectivo en laboratorio de uno decepcionante en la clínica.
Señalización GPCR canónica en CB1 y CB2
Secuencia de señalización canónica de CB1/CB2
- Unión del ligando Un agonista estabiliza una conformación activa del receptor.
- Activación de la proteína G El receptor favorece el intercambio de GDP por GTP en Gi/o.
- Separación de subunidades Galpha y Gbeta-gamma regulan efectores aguas abajo.
- Cambio del segundo mensajero La actividad de la adenilil ciclasa disminuye y el cAMP desciende.
- Efecto celular Cambian los canales iónicos, la liberación de neurotransmisores, las quinasas y la regulación génica.
En el esquema canónico, la unión del agonista estabiliza una conformación activa del receptor, el receptor actúa como factor de intercambio de nucleótidos de guanina para Gi/o, Gαi/o intercambia GDP por GTP, y los componentes Gα y Gβγ regulan entonces efectores descendentes. Para CB1 y CB2, la lectura clásica es la inhibición de la adenilil ciclasa y una disminución del AMPc intracelular. Ese hallazgo se convirtió en una de las primeras firmas bioquímicas utilizadas para definir la actividad de los receptores cannabinoides.
Pero “canónico” no debe leerse como “uniforme”. CB1 muestra una alta actividad constitutiva en varios sistemas de expresión, lo que significa que el receptor puede señalar de forma medible incluso en ausencia de agonista. Esa propiedad ayuda a explicar por qué los inversos agonistas como rimonabant hicieron más que bloquear el tono cannabinoide endógeno; empujaron la señalización por debajo de la línea basal y produjeron efectos adversos centrales marcados. CB2 también se acopla a Gi/o, pero la forma en que los ligandos estabilizan estados activos difiere de CB1. Un trabajo estructural revisado en Frontiers in Chemical Biology en 2026 subrayó que la selectividad de subtipo entre “CB1 y CB2” está impulsada por diferencias a nivel del receptor que alteran no solo la unión, sino también la eficacia y la regulación. Un estudio indexado en PubMed de 2025/2026 sobre selectividad de subtipo fue más allá al sostener que la selectividad de los endocannabinoid es dinámica, moldeada por el comportamiento conformacional más que por un modelo fijo de llave-cerradura.
Esta es una de las razones por las que los fitocannabinoides, endocannabinoid y ligandos sintéticos nunca deben tratarse como intercambiables. La anandamida, identificada por Raphael Mechoulam y Lumír Hanuš, y el 2-araquidonoilglicerol son ligandos endógenos producidos a demanda e inactivados rápidamente. Δ9-THC es un agonista parcial de origen vegetal con cinética y eficacia distintas de las de esos endocannabinoid. Agonistas sintéticos como CP55,940, WIN55,212-2 o HU-210 suelen impulsar una activación más intensa del receptor y pueden reclutar vías de señalización en distintos grados. Algunos ligandos favorecen la señalización por proteínas G sobre el reclutamiento de β-arrestina; otros no. El artículo de American Journal of Psychiatry en 2025 hizo ese punto directamente para CB1, argumentando que la señalización sesgada es una estrategia terapéutica plausible en la esquizofrenia, un trastorno que afecta a unos 24 millones de personas en todo el mundo.
CB2 añade otra corrección a simplificaciones anteriores. Sigue estando enriquecido en muchas poblaciones de células inmunitarias, pero la revisión de 2026 de Frontiers in Behavioral Neuroscience describió “una actualización de los últimos 3 años” en la que la señalización de CB2 ganó atención en trastornos del sistema nervioso central vinculados con neuroinflamación y neurodegeneración. Así pues, incluso antes de introducir el agonismo sesgado, la antigua dicotomía de un “receptor cerebral” y un “receptor inmunitario” ya falla a nivel de contexto de señalización.
Efectos sobre AMPc, canales iónicos y liberación de neurotransmisores
El efecto de segundo mensajero mejor conocido en ambos receptores es la supresión de la formación de AMPc mediante la inhibición de la adenilil ciclasa. Menor AMPc suele significar menor actividad de la proteína quinasa A, fosforilación alterada de dianas descendentes y cambios más lentos en la expresión génica a través de vías como CREB. En las neuronas, sin embargo, los efectos rápidos suelen ser más importantes que los lentos.
CB1 está fuertemente posicionado para el control presináptico. En muchos circuitos cerebrales se localiza en terminales axónicas, donde la activación del receptor reduce la probabilidad de liberación de neurotransmisor. Esto ocurre mediante una combinación de inhibición mediada por Gβγ de los canales de calcio dependientes de voltaje y activación de conductancias de potasio rectificadoras entrantes u otras corrientes de potasio que amortiguan la excitabilidad de la terminal. Menor entrada de calcio significa menos vesículas fusionadas. El resultado es menos neurotransmisor liberado en la hendidura sináptica.
Ese es el mecanismo central de la señalización endocannabinoid retrógrada de corto alcance. Una neurona postsináptica se activa, sintetiza endocannabinoid a demanda a partir de precursores lipídicos de membrana y los envía hacia atrás a través de la sinapsis para activar los receptores CB1 presinápticos. La terminal presináptica entonces libera menos neurotransmisor. Es un freno de retroalimentación. En sinapsis excitadoras, eso puede suprimir la liberación de glutamato; en sinapsis inhibidoras, puede suprimir la liberación de GABA. La dirección de la salida del circuito depende de qué terminal exprese CB1. El mismo receptor, consecuencias opuestas en la red.
| Término | Qué se suprime | Mecanismo descrito |
|---|---|---|
| DSI | Inhibición | La actividad postsináptica libera endocannabinoides que activan CB1 presináptico y reducen la liberación de GABA |
| DSE | Excitación | La actividad postsináptica libera endocannabinoides que activan CB1 presináptico y reducen la liberación de glutamato |
DSI y DSE Formas a corto plazo de plasticidad sináptica mediada por endocannabinoides en las que la despolarización postsináptica suprime la transmisión inhibidora (DSI) o la transmisión excitadora (DSE) mediante la activación presináptica de CB1.
Los términos fisiológicos clásicos lo capturan: supresión de la inhibición inducida por despolarización, DSI, y supresión de la excitación inducida por despolarización, DSE. Ambas son formas de plasticidad sináptica a corto plazo impulsadas por la liberación de endocannabinoid y la activación presináptica de CB1. También ocurren efectos de mayor duración, incluida la depresión a largo plazo mediada por endocannabinoid en algunas sinapsis. Esos fenómenos importan porque conectan la bioquímica del receptor con la conducta: el procesamiento del dolor, la extinción del miedo, el aprendizaje de hábitos, el apetito, el control motor y el umbral convulsivo dependen todos de este ajuste de la probabilidad de liberación.
Los detalles no son triviales. Un agonista parcial como Δ9-THC puede no reproducir el patrón completo producido por una breve ráfaga endógena de 2-AG. Tampoco un agonista sintético completo preservará necesariamente la temporalidad fisiológica. La dosis importa. También importa la reserva de receptores. En una sinapsis con una expresión densa de CB1, incluso un agonista parcial puede producir un gran efecto sobre la liberación de neurotransmisor. En un tejido diferente, el mismo ligando puede parecer débil.
CB2 tiene una fisiología sináptica directa menos establecida que CB1, pero también reduce el AMPc y puede regular la señalización del calcio, las vías de quinasas y la liberación de mediadores inflamatorios en células inmunitarias y gliales. Eso hace que CB2 sea relevante para la comunicación neurona-glía, especialmente en estados de enfermedad en los que cambia la expresión del receptor. El artículo de análisis de redes indexado en PubMed en 2025/2026 trató CB1 y CB2 como nodos influyentes en una señalización más amplia de endocannabinoid y metabolismo, lo cual es un marco mejor que tratarlos como interruptores aislados.
Desensibilización, internalización y regulación del receptor
Cómo se adaptan los receptores a la exposición repetida a agonistas
- Fosforilación Las regiones intracelulares del receptor son modificadas por quinasas del GPCR y otras quinasas.
- Reclutamiento de beta-arrestina Las arrestinas desacoplan los receptores de las proteínas G y pueden iniciar señalización adicional.
- Desensibilización El receptor se vuelve menos sensible.
- Internalización Los receptores son incorporados a vías endocíticas.
- Destino tras la captación Los receptores pueden reciclarse de vuelta a la superficie o degradarse.
Ningún receptor puede activarse de forma continua sin consecuencias. Para CB1 y CB2, la exposición prolongada o repetida a agonistas conduce comúnmente a la fosforilación de las regiones intracelulares del receptor por quinasas GRK y otras quinasas, al reclutamiento de β-arrestinas, al desacoplamiento de las proteínas G y luego a la internalización a través de vías endocíticas. La desensibilización ocurre primero. A menudo le sigue la endocitosis. Después viene el reciclaje o la degradación.
Para CB1, este ciclo regulador es una de las principales razones por las que los efectos agudos y crónicos difieren. Los agonistas potentes pueden desencadenar una desensibilización rápida en sistemas celulares y tolerancia medible in vivo. La regulación específica por región importa aquí. Los receptores CB1 no se desensibilizan por igual en todas las poblaciones neuronales, lo que ayuda a explicar por qué la tolerancia se desarrolla de manera desigual entre los efectos cannabinoides. Las respuestas analgésicas, la hipotermia, la alteración de la memoria y los efectos motores pueden cambiar a ritmos distintos porque el receptor se regula de manera diferente en distintos circuitos.
Las β-arrestinas no son solo interruptores de apagado. También pueden actuar como andamios para sus propias cascadas de señalización, incluidas las vías de MAP quinasa, razón por la cual el reclutamiento de arrestina se ha vuelto central para el agonismo sesgado. Un ligando que inhibe fuertemente el AMPc pero recluta débilmente β-arrestina puede comportarse de manera distinta de uno que hace ambas cosas con eficiencia. Eso ya no es un matiz teórico; es una estrategia de diseño de fármacos. La discusión de 2025 en American Journal of Psychiatry sobre la sesgo de CB1 en la esquizofrenia refleja una lección más amplia de los GPCR: evitar ciertas ramas de señalización puede reducir algunas desventajas, pero la selectividad por una vía no garantiza el éxito clínico.
La internalización en sí también depende del ligando. Algunos agonistas impulsan una endocitosis extensa del receptor; otros producen una internalización limitada a pesar de la activación de proteínas G. Los moduladores alostéricos complican aún más el panorama al cambiar cómo los ligandos ortostéricos estabilizan los estados del receptor. Aquí es donde la farmacología estructural se encuentra con la terapéutica. La revisión estructural de 2026 dejó claro que la conformación del receptor controla juntos la eficacia de señalización y la regulación del receptor, no como temas separados.
Esa es la lección clave de señalización que hay que conservar. CB1 y CB2 no son detectores simples de encendido y apagado para los cannabinoides. Son nodos regulados cuyo resultado cambia a lo largo de milisegundos a días. Cualquier intento serio de dirigirlos, ya sea para epilepsia, dolor, psicosis o enfermedad inflamatoria, tiene que tener en cuenta el acoplamiento Gi/o, los segundos mensajeros, el control de canales iónicos, la localización sináptica y el hecho de que el receptor se adaptará a la estimulación.
Señalización sesgada: por qué un receptor puede producir diferentes resultados biológicos
La antigua visión de la farmacología del cannabinoid trataba a un receptor como si fuera un interruptor de la luz: los agonistas lo activan, los antagonistas lo apagan, y todo lo demás depende de dónde se exprese ese receptor. Esa imagen no es adecuada para CB1 o CB2. No explica por qué dos ligandos que actúan en el mismo receptor pueden producir efectos conductuales, cognitivos, inflamatorios o terapéuticos marcadamente diferentes. Tampoco explica por qué el descubrimiento de fármacos dirigidos a los cannabinoid receptors ha producido repetidamente compuestos que parecían prometedores in vitro pero resultaron decepcionantes, intolerables o clínicamente ambiguos.
Eso importa mucho más allá de la teoría académica de los receptores. La World Health Organization estimó que 200 millones de personas usaron cannabis en 2019, alrededor del 4% de la población mundial de 15 a 64 años. La esquizofrenia afecta a unas 24 millones de personas en todo el mundo. En ese contexto, la farmacología de CB1 no es un asunto marginal. Se sitúa en la intersección de la salud pública, la psiquiatría y el diseño de fármacos. La U.S. FDA, a fecha de 2025, había aprobado un producto farmacéutico derivado de cannabis y tres productos farmacéuticos relacionados con cannabis, una cifra pequeña en comparación con la magnitud del interés clínico. Una razón por la que el progreso ha sido más lento de lo que sugiere el debate público es que la señalización de los cannabinoid receptors no consiste simplemente en ocupación del receptor. Consiste en selección de vía.
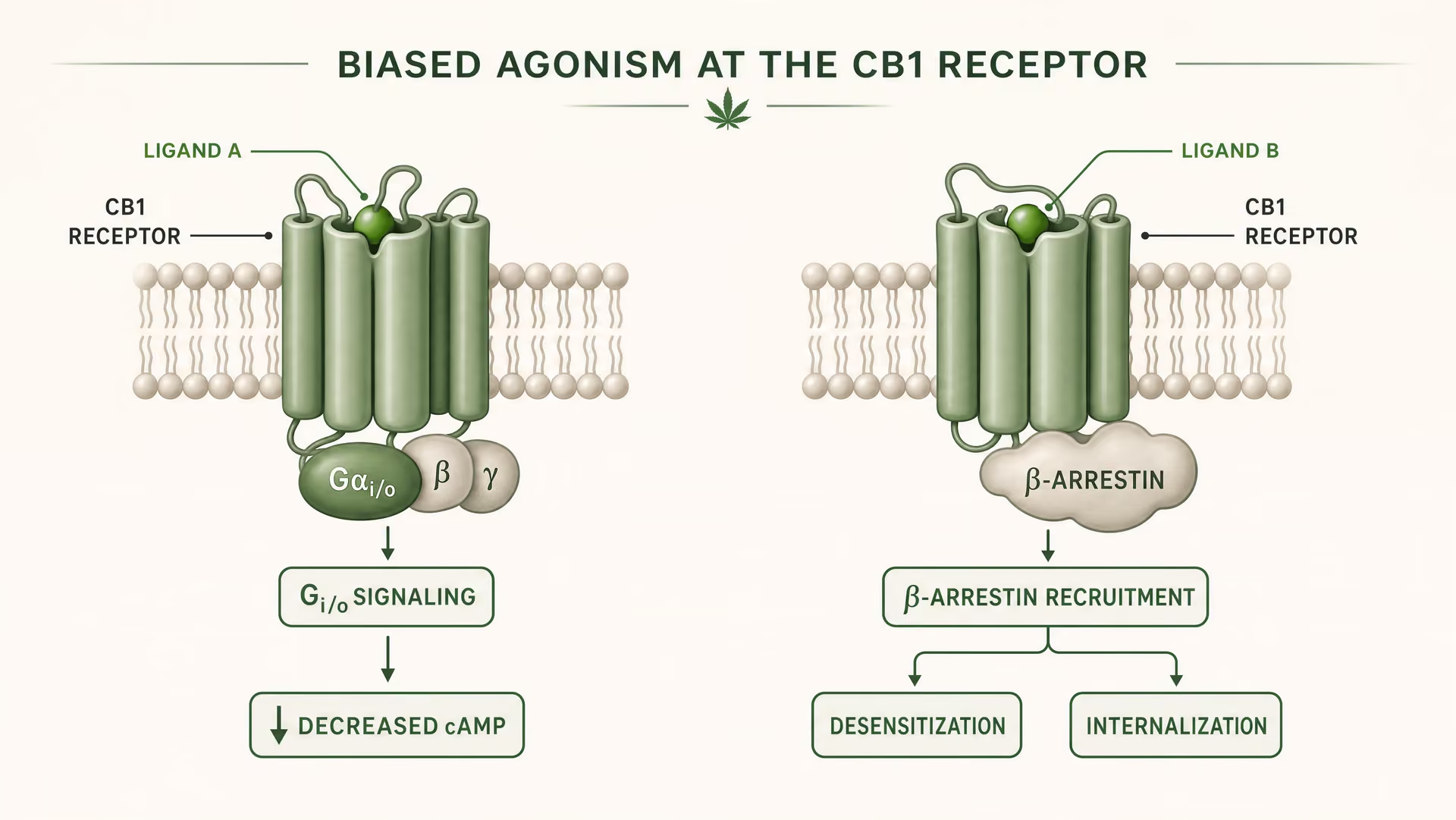
Qué significa el agonismo sesgado en la farmacología de los GPCR
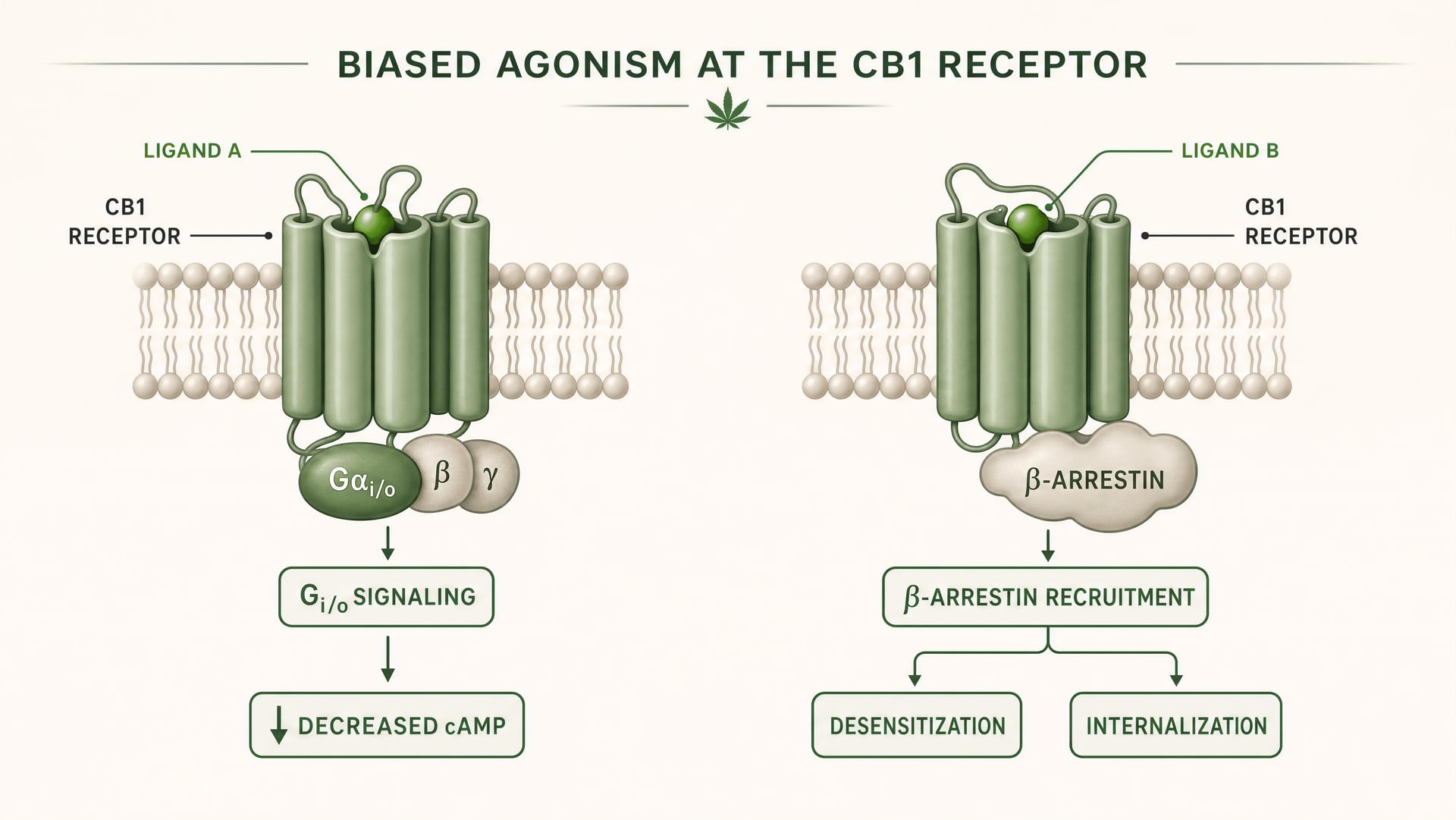
CB1 y CB2 son receptores acoplados a proteínas G de clase A. El trabajo fundacional de Allyn Howlett estableció CB1 como un cannabinoid receptor acoplado a Gi/o, ayudando a mover el campo desde una farmacología imprecisa hacia mecanismos definidos por receptores. Pero el acoplamiento a Gi/o es solo el comienzo de la historia. Una vez que un ligando se une, el receptor puede adoptar más de una forma activa, y esas formas no señalan de manera idéntica. Algunas conformaciones del receptor favorecen la activación de la proteína G. Otras reclutan con mayor fuerza a beta-arrestins. Algunos estados promueven la fosforilación, la desensibilización o la internalización del receptor. Otros producen una señalización más duradera desde la membrana plasmática o desde compartimentos endosómicos.
{kind=link}
Agonismo sesgado Propiedad de un ligando que favorece algunas vías de señalización aguas abajo sobre otras en el mismo receptor.
Eso es agonismo sesgado en términos simples: distintos ligandos estabilizan distintas conformaciones del receptor, y esas conformaciones favorecen distintas vías posteriores. Un receptor no está simplemente encendido o apagado. Está dirigido conformacionalmente.
Para CB1, esto es especialmente importante porque el receptor se encuentra en un entorno de señalización denso, plástico y muy dependiente del tipo celular. En una terminal glutamatérgica cortical, un ligando puede reducir la liberación de neurotransmisor mediante la inhibición de la adenylyl cyclase mediada por Gi/o y la modulación de canales iónicos. En una interneurona GABAérgica, el mismo receptor puede desplazar el equilibrio del circuito local en una dirección muy distinta. Si además el ligando promueve un reclutamiento intenso de beta-arrestins, el receptor puede internalizarse más rápido, acortando un efecto mientras abre otro. Cambia la cronología. Cambia la ubicación de la señal. Cambia la respuesta fisiológica.
Esto no es una sutileza teórica. La revisión estructural de 2026 en Frontiers in Chemical Biology sobre cannabinoid receptors deja claro el punto: la selectividad de ligando en CB1 y CB2 depende de diferencias estructurales a nivel del receptor que alteran la unión, la eficacia de la señalización y la regulación del receptor. La palabra clave ahí es regulación. Un ligando puede ser similar en afinidad y, sin embargo, diferir en eficacia, reclutamiento de arrestina, tiempo de residencia o propensión a desencadenar desensibilización. El estudio indexado en PubMed de 2025/2026 sobre el mecanismo dinámico de la selectividad de subtipos lleva la misma idea más lejos al sostener que la selectividad emerge de la dinámica conformacional, no de un modelo estático de llave y cerradura. Por tanto, los endocannabinoids, los phytocannabinoids y los ligandos sintéticos no deben agruparse en una misma categoría. Anandamide, descubierto por Raphael Mechoulam y Lumír Hanuš, no se comporta como delta-9-tetrahydrocannabinol, y ninguno de los dos se comporta como una sonda sintética altamente optimizada.
La señalización sesgada también explica por qué los moduladores alostéricos atraen tanto interés. Un ligando alostérico puede no activar directamente CB1 como lo hace un agonista ortostérico, y aun así puede remodelar las preferencias de señalización del receptor, amplificando una vía y atenuando otra. Eso abre una vía hacia un control fino. En principio.
La señalización sesgada de CB1 como línea de investigación en esquizofrenia
El artículo de 2025 en el American Journal of Psychiatry presenta el argumento más sólido de los últimos años de que la señalización sesgada de CB1 no es solo un concepto de farmacología, sino una estrategia terapéutica plausible para la esquizofrenia. Ese argumento merece atención porque la investigación sobre esquizofrenia ha abordado tradicionalmente los cannabinoids desde la epidemiología, la asociación con el riesgo o advertencias amplias sobre psicosis. El artículo de AJP cambia el marco. Pregunta si el problema no son los “cannabinoids” en general, ni siquiera la “activación de CB1” en general, sino qué estados de señalización de CB1 se están activando, en qué circuitos y durante cuánto tiempo.
Esa es una pregunta mucho mejor.
CB1 es uno de los GPCR más abundantes del cerebro, con una expresión elevada en la corteza, el hipocampo, los ganglios basales y el cerebelo, pero la abundancia por sí sola no explica los efectos clínicos. La esquizofrenia implica una desregulación de la relevancia, la cognición, la percepción y la coordinación de redes a través de sistemas corticales y subcorticales. Un receptor situado para modular la actividad de circuitos relacionados con glutamato, GABA y dopamina es, por tanto, relevante por diseño. El artículo de AJP sostiene que los ligandos sesgados de CB1 podrían separar los efectos útiles sobre los circuitos de las limitaciones como la alteración cognitiva, la ansiedad, la disforia o las respuestas psicotomiméticas.
Es una afirmación ambiciosa, pero no una especulación vacía. Sigue la lógica más amplia del campo de los GPCR, donde el sesgo de vía ya ha cambiado la manera en que los investigadores piensan sobre fármacos opioides, angiotensina y receptores de dopamina. La esperanza traslacional en CB1 es que determinadas salidas de señalización puedan mejorar la función de las redes corticales o amortiguar estados aberrantes del circuito sin reproducir el perfil completo de efectos adversos asociado con el agonismo de alta eficacia de CB1.
La esquizofrenia es un buen caso de prueba porque el listón clínico es alto. Un candidato a fármaco no puede limitarse a alterar el comportamiento en un ensayo con roedores. Debe evitar empeorar la psicosis, la sedación y la alteración cognitiva en personas ya vulnerables a esos problemas. Eso convierte de inmediato la selectividad de vía en algo más que una preferencia de química medicinal. La transforma en un requisito de seguridad.
El marco de AJP también ayuda a corregir una simplificación común en las discusiones sobre cannabis. Delta-9-THC es un phytocannabinoid con actividad agonista parcial en CB1, pero sus efectos reflejan la dosis, el momento, la reserva de receptor, el tono endocannabinoid local y el compromiso de vías en distintas poblaciones neuronales. Un ligando sintético de CB1 diseñado para favorecer una ruta intracelular concreta podría comportarse de manera muy distinta a THC aunque ambos “actúen sobre CB1”. Lo contrario también es cierto: dos compuestos que mejoran un desenlace preclínico relevante para la esquizofrenia podrían divergir mucho en cognición o afecto si uno impulsa una señalización dominada por arrestina y el otro no. La identidad del receptor por sí sola no puede predecir todo el fenotipo.
Por qué la selectividad de vía importa para la seguridad y la eficacia
La selectividad de vía importa porque la eficacia no es una dimensión única. Un fármaco cannabinoid puede ser potente y aun así clínicamente pobre. Puede ser selectivo para CB1 y aun así fracasar. Puede evitar CB2 por completo y aun así producir efectos inmunitarios o metabólicos no deseados mediante interacciones en red. El análisis integrador de redes indexado en PubMed de 2025/2026 identificó a CB1 y CB2 como nodos altamente influyentes en el endocannabinoid system y mapeó su señalización sobre vías metabólicas. Esa visión sistémica es esencial. Los receptores no operan de forma aislada, y el sesgo de vía en un nodo puede propagarse hacia programas fisiológicos más amplios.
Para CB1, las preocupaciones de seguridad son obvias. Una activación central intensa de CB1 puede producir deterioro de la memoria, alteración de la percepción, ansiedad, taquicardia y, en individuos susceptibles, efectos relacionados con la psicosis. Cualquier programa terapéutico orientado al dolor, el apetito, el estado de ánimo, la adicción o la esquizofrenia debe afrontar ese perfil de riesgo. Un ligando que conserve un efecto sináptico deseado mediado por Gi/o mientras limita la desensibilización impulsada por beta-arrestin u otras cascadas de señalización adversas podría, en teoría, ampliar la ventana terapéutica. Pero “en teoría” importa. Muchos programas de ligandos sesgados en la farmacología de GPCR han mostrado que el sesgo medido en un sistema de ensayo no siempre predice los resultados in vivo. El contexto celular, la densidad del receptor, la expresión del efector y la cinética pueden modificar por completo el sesgo aparente.
CB2 ofrece un paralelo de advertencia. La revisión de 2026 en Frontiers in Behavioral Neuroscience describe una actualización de los últimos 3 años en la que la señalización de CB2 ganó atención en trastornos del sistema nervioso central por sus vínculos con mecanismos neuroinflamatorios y neurodegenerativos. Eso socava directamente la vieja idea de que CB2 es irrelevante para el cerebro. Aun así, simplemente dirigir la acción hacia CB2 no garantiza un medicamento antiinflamatorio útil. La distribución es más matizada de lo que sugería la antigua división entre cerebro y cuerpo, y las consecuencias de la señalización siguen dependiendo del ligando y del contexto.
Por tanto, la lección práctica es clara: la selectividad por subtipo de receptor es necesaria, pero no suficiente. La selectividad de vía puede ser la diferencia entre un cannabinoid que parece terapéutico, uno que es intoxicante y otro que fracasa en los ensayos porque no logra separar el beneficio del efecto adverso. Para CB1, especialmente en psiquiatría, esa distinción probablemente decidirá si el receptor sigue siendo una advertencia o se convierte en un objetivo farmacológico viable.
Biología estructural de CB1 y CB2: cómo la forma impulsa la selectividad
La biología estructural cambió la manera en que se discuten los receptores de cannabis. La antigua simplificación —CB1 explica la intoxicación, CB2 explica la inflamación— pasa por alto el hecho de que ambos receptores son receptores acoplados a proteína G de clase A, cuyo comportamiento depende de la forma, el movimiento, la profundidad de unión y los socios de señalización disponibles en una célula determinada. Esto importa mucho más allá de la farmacología básica. Se estima que cannabis fue utilizado por 200 millones de personas en 2019, o el 4 % de la población mundial de 15 a 64 años, según la OMS, pero la FDA todavía solo enumera un producto farmacéutico derivado de cannabis y tres productos farmacéuticos relacionados con cannabis como aprobados en 2025. Una de las razones de esa brecha es estructural: es difícil diseñar ligandos cannabinoid que actúen sobre el receptor correcto, de la manera correcta, durante la duración correcta.
La revisión de Frontiers in Chemical Biology de 2026 deja este punto claro. CB1 y CB2 no solo difieren en dónde se expresan. También difieren en la arquitectura de sus cavidades de unión de ligando, en la forma y flexibilidad de sus bucles extracelulares, en el empaquetamiento de sus hélices transmembrana y en los estados conformacionales que prefieren después de que un ligando se une. Esas características influyen no solo en la selectividad, sino también en la eficacia, la desensibilización, la internalización y el sesgo de vía.
Qué revelan los estudios estructurales sobre los bolsillos del receptor
Una cavidad ortostérica es la principal cavidad de unión donde ligandos endógenos como la anandamida y el 2-arachidonoylglycerol, fitocannabinoides como THC y muchos ligandos sintéticos realizan su contacto primario. En CB1 y CB2, esa cavidad se sitúa dentro del haz de siete hélices transmembrana, parcialmente cubierta por bucles extracelulares que pueden abrir el acceso o restringirlo.
Las estructuras de cryo-EM y de rayos X de los últimos años mostraron que los receptores de cannabis no se comportan como cerraduras rígidas que esperan una llave. Se comprenden mejor como blancos móviles con formas preferidas. La revisión de Frontiers in Chemical Biology de 2026 subraya que las cavidades ortostéricas de CB1 y CB2 son lo bastante similares como para unir clases de ligandos superpuestas, pero lo bastante distintas en tamaño, identidad de residuos y flexibilidad local como para desplazar la afinidad y el resultado de señalización. Por eso compuestos estrechamente relacionados pueden separarse farmacológicamente. Un pequeño cambio en el volumen del sustituyente, la polaridad o la longitud de la cadena puede alterar hasta qué profundidad penetra un ligando en la cavidad, qué hélices empuja y si el receptor se estabiliza en un estado que favorece a la proteína G o a la arrestina.
CB1 ha sido especialmente informativo desde el punto de vista estructural porque ahora existen muchos modelos de alta resolución de estados inactivos y activos. Un tema recurrente es que su cavidad es amplia e hidrofóbica, lo que se ajusta a la naturaleza lipofílica de muchos cannabinoid. El bucle extracelular 2 y las partes superiores de varias hélices ayudan a dar forma al acceso. Las hélices transmembrana son los siete segmentos que atraviesan la membrana y constituyen el núcleo del receptor; cuando un ligando se une, estas hélices pueden desplazarse entre sí. El movimiento más importante desde el punto de vista farmacológico suele producirse en el lado intracelular, donde el movimiento hacia afuera de la hélice 6 ayuda a crear un sitio de unión para las proteínas Gi/o. Ese desplazamiento es uno de los rasgos distintivos de la activación del receptor.
CB2 comparte el mismo pliegue general de GPCR, pero la revisión de Frontiers sostiene que las diferencias específicas de subtipo en los aminoácidos alrededor de la cavidad y las regiones de bucle proporcionan a los químicos medicinales puntos de apoyo explotables para la selectividad. La idea no es que una cavidad sea simplemente “similar al cerebro” y la otra “similar al sistema inmunitario”. La idea es geométrica y energética. Residuos distintos alteran el contorno de la cavidad, las opciones locales de enlace por hidrógeno, el apilamiento aromático y la flexibilidad de los canales de acceso por los que los ligandos entran desde la membrana.
Un estudio indexado en PubMed de 2025/2026 sobre el mecanismo dinámico de la selectividad de subtipo llevó esto más lejos al argumentar que la selectividad de endocannabinoid no es solo una cuestión de afinidad de unión estática. La dinámica conformacional importa. En términos simples, un receptor puede muestrear múltiples formas antes y después de la unión del ligando, y algunos ligandos estabilizan una forma selectiva mejor que otros. Esto ayuda a explicar por qué los lípidos endógenos, los fitocannabinoides y los ligandos sintéticos pueden mostrar preferencias de subtipo distintas, incluso cuando sus esqueletos parecen relacionados sobre el papel.
Determinantes de la selectividad de ligandos entre CB1 y CB2
La selectividad comienza con la química de contacto, pero no termina ahí. La revisión de Frontiers in Chemical Biology presenta la selectividad como el producto de diferencias estructurales a nivel del receptor que afectan simultáneamente la unión, la eficacia de señalización y la regulación. Ese es el marco correcto. Un ligando puede ser selectivo para CB2 en un ensayo de unión de radioligando y, aun así, perder su ventaja práctica si también impulsa estados del receptor que causan tolerancia rápida o señalización débil en células relevantes para la enfermedad.
Varias características estructurales aparecen una y otra vez. En primer lugar, la composición de aminoácidos de la cavidad ortostérica difiere lo suficiente entre CB1 y CB2 como para cambiar cómo se acomodan el grupo cabeza, el núcleo y la cadena hidrofóbica de un ligando. En segundo lugar, los bucles extracelulares ayudan a dar forma a la entrada y la orientación. En tercer lugar, las regiones superiores y medias de las hélices transmembrana pueden sesgar al receptor hacia conjuntos de estados activos ligeramente distintos. Un estado conformacional es simplemente una de las formas posibles del receptor en un momento dado. Los distintos ligandos no solo se unen a un receptor; estabilizan un subconjunto de esas formas.
Por eso la selectividad de subtipo suele ser modesta para los cannabinoid naturales. THC, por ejemplo, interactúa con ambos receptores. Anandamida y 2-AG también actúan sobre ambos, aunque con diferencias dependientes del contexto en potencia, eficacia y metabolismo. Los ligandos sintéticos han sido más útiles para separar las relaciones estructura-selectividad porque los químicos pueden alterar de forma sistemática características como la longitud de la cadena lateral, las restricciones del anillo y los sustituyentes polares. Incluso así, una separación limpia es difícil. CB1 y CB2 son lo bastante homólogos como para que un compuesto diseñado para uno a menudo conserve una actividad significativa sobre el otro.
Esto tiene consecuencias prácticas. Durante mucho tiempo, los desarrolladores de fármacos han buscado agonistas selectivos de CB2 con la esperanza de obtener efectos antiinflamatorios o analgésicos sin los efectos adversos centrales intensos mediados por CB1. A veces esa estrategia funciona farmacológicamente, pero no es un pase libre. La revisión de Frontiers in Behavioral Neuroscience de 2026 subraya que CB2 ha atraído atención en trastornos del sistema nervioso central durante “los últimos 3 años”, lo que debilita la visión simple de que CB2 es irrelevante para el cerebro. Por tanto, incluso un ligando de CB2 “periférico” no puede interpretarse con mapas de receptores obsoletos.
Por qué la eficacia y la regulación también son cuestiones estructurales
La potencia responde cuánta cantidad de ligando se necesita. La eficacia pregunta qué hace ese ligando al receptor una vez unido. La biología estructural vincula ambas cosas, pero no las confunde. Dos ligandos pueden ocupar la misma cavidad y producir respuestas muy diferentes porque estabilizan distintas conformaciones activas.
Para CB1, esto es central para el pensamiento terapéutico actual. El artículo de American Journal of Psychiatry de 2025 sostiene que la señalización sesgada en CB1 puede ser una estrategia plausible para la esquizofrenia, un trastorno que afecta a unos 24 millones de personas en todo el mundo. El agonismo sesgado significa que un ligando favorece una vía descendente sobre otra, a menudo la señalización Gi/o frente al reclutamiento de beta-arrestina, o viceversa. Estructuralmente, ese sesgo surge porque el ligando desplaza la cara intracelular del receptor hacia una forma que encaja mejor con un socio de señalización que con otro. No es una idea abstracta. Es un objetivo de química medicinal.
Las beta-arrestinas son importantes porque contribuyen a la desensibilización y la internalización. La desensibilización significa que el receptor responde menos después de una activación repetida o sostenida. La internalización significa que el receptor es retirado de la membrana de la superficie hacia el interior de la célula. Ambos procesos están influidos por la conformación del receptor. Algunos ligandos activan con fuerza las proteínas G pero reclutan débilmente arrestinas; otros hacen ambas cosas. Un fármaco de CB1 tolerable desde el punto de vista conductual puede necesitar precisamente este tipo de separación, en lugar de un bloqueo simple o un agonismo completo.
La modulación alostérica añade otra capa. Un ligando alostérico se une fuera de la cavidad ortostérica y cambia la respuesta del receptor a otros ligandos. Estructuralmente, esto resulta atractivo porque podría permitir un control más fino de la forma del receptor que golpear la cavidad principal con un agonista de alta eficacia. Para la farmacología de cannabis, eso podría significar preservar la señalización útil mientras se reducen los efectos adversos vinculados a una activación excesiva de CB1.
La lección más amplia es contundente. La forma del receptor no es un detalle secundario. Es la explicación de por qué los fitocannabinoides, los endocannabinoid y los compuestos sintéticos pueden producir perfiles clínicos distintos incluso cuando se dirigen a la misma familia de receptores. Dado que CB1 y CB2 ocupan nodos influyentes en los análisis de red del sistema endocannabinoid, como informó un estudio de sistemas indexado en PubMed de 2025/2026, los errores en selectividad o eficacia pueden propagarse por múltiples vías. La información estructural no garantiza un fármaco exitoso. Sí explica por qué sigue siendo tan difícil alinear selectividad, potencia y tolerabilidad.
Endocannabinoids y selectividad de subtipo: anandamida, 2-AG y comportamiento dinámico de los receptores
La farmacología de los endocannabinoids suele explicarse con una simplificación que suena ordenada, pero resulta engañosa: la anandamida y el 2-arachidonoylglycerol (2-AG) se tratan como si fueran los “agonistas estándar” incorporados del organismo, útiles principalmente como puntos de referencia para lo que CB1 y CB2 hacen normalmente. Ese enfoque pasa por alto el hecho central. No se trata de hormonas estables que circulen libremente y encuentren receptores en condiciones uniformes. Son mediadores lipídicos de vida corta, producidos a demanda en las membranas por sistemas enzimáticos específicos de cada célula, y luego destruidos con rapidez. Su aparente preferencia por un receptor puede cambiar según la composición de la membrana, las conformaciones del receptor, la actividad enzimática local y el momento de la liberación.
Eso importa más allá de la teoría académica sobre receptores. Según la WHO, en 2019 se estimó que cannabis fue utilizado por 200 millones de personas, es decir, el 4% de la población mundial de 15 a 64 años, y los medicamentos dirigidos a cannabinoid ya están en la clínica: la FDA afirmó en 2025 que ha aprobado un producto farmacológico derivado de cannabis y tres productos farmacológicos relacionados con cannabis. Si los ligandos endógenos no se comportan como controles sencillos de libro de texto, entonces la comparación de fitocannabinoids o candidatos a fármacos con ellos no es trivial.
Por qué los ligandos endógenos no son compuestos de referencia simples
La anandamida, identificada en 1992 por Devane, Hanus, Breuer, Mechoulam y colegas, y el 2-AG, establecido unos años después como otro endocannabinoid importante, difieren en algo más que en tablas de potencia. Proceden de rutas biosintéticas distintas, alcanzan los receptores en escalas temporales diferentes y son eliminados por enzimas diferentes. La anandamida se genera principalmente a partir de N-arachidonoyl phosphatidylethanolamine mediante vías que implican NAPE-PLD, aunque existen rutas alternativas. El 2-AG se produce en gran medida a partir de diacylglycerol por acción de DAGLα y DAGLβ. No son notas bioquímicas menores; determinan dónde y cuándo ocurre la activación del receptor.
Incluso la afirmación más antigua de que la anandamida es un ligando con preferencia por CB1, mientras que el 2-AG es un agonista completo tanto en CB1 como en CB2, puede volverse demasiado burda cuando se traslada de sistemas purificados a tejido vivo. La anandamida también es sustrato de fatty acid amide hydrolase, o FAAH, y en ciertas condiciones puede interactuar con dianas no cannabinoid, como TRPV1. En muchas mediciones, el 2-AG está presente en tejido cerebral a niveles basales mucho más altos que la anandamida y se hidroliza principalmente por monoacylglycerol lipase, o MAGL, con ABHD6 y ABHD12 también contribuyendo. Por tanto, la “entrada” del receptor que proporciona cada ligando no depende solo de la afinidad o la eficacia en CB1 frente a CB2. Depende de si el ligando se produce en la sinapsis, dentro de una célula inmunitaria, cerca de una membrana intracelular o en un compartimento rico en enzimas degradativas.
El trabajo de farmacología de receptores de Allyn Howlett ayudó a establecer CB1 como un auténtico GPCR acoplado en gran medida a proteínas Gi/o. Ese marco sigue siendo esencial, pero ya no sostiene un modelo simple de ligando endógeno como fármaco de referencia. CB1 y CB2 pueden señalizar mediante inhibición de adenylate cyclase, regulación de canales iónicos, vías de MAP kinase, reclutamiento de beta-arrestin, internalización del receptor y desensibilización. Un ligando que parece “más débil” en un ensayo puede generar una señalización más prolongada en un tejido donde la degradación es lenta, o puede favorecer una vía downstream sobre otra. El artículo de 2025 en American Journal of Psychiatry sobre biased signaling de CB1 subraya este punto en un contexto patológico: en trastornos como la schizophrenia, que afecta a unas 24 millones de personas en todo el mundo, la cuestión terapéutica no es simplemente si se activa CB1, sino cómo.
Mecanismos dinámicos de la selectividad de subtipo
El reciente estudio sobre selectividad de subtipo indexado en PubMed (PMID: 41962866) es importante porque aleja al campo de una historia de llave y cerradura. Su mensaje central es que la selectividad de los endocannabinoids puede surgir de mecanismos dinámicos. En otras palabras, la preferencia por un subtipo de receptor no se explica solo por una diferencia fija en la afinidad de unión de un ligando a CB1 frente a CB2. El complejo ligando-receptor explora múltiples estados conformacionales, y esos estados difieren entre CB1 y CB2.
Ese hallazgo encaja con la revisión de 2026 en Frontiers in Chemical Biology, que sostiene que las diferencias estructurales entre CB1 y CB2 moldean la unión, la eficacia de señalización y la regulación del receptor. Los receptores comparten una similitud de secuencia considerable, pero pequeñas diferencias en la arquitectura del bolsillo ortostérico, los bucles extracelulares, los movimientos de las hélices transmembrana y las superficies de acoplamiento intracelular pueden sesgar lo que ocurre tras la unión del ligando. Un ligando puede entrar en bolsillos similares en ambos receptores, pero estabilizar un estado más competente para señalizar en un subtipo, o favorecer transiciones distintas entre estados inactivo, intermedio y activo.
En el caso de los endocannabinoids, esto es especialmente plausible porque son lípidos flexibles y no andamios sintéticos rígidos. La flexibilidad es una variable farmacológica. La anandamida puede adoptar múltiples conformaciones, y el 2-AG es igualmente dinámico en las membranas. La cuestión entonces no es solo “¿se une?”, sino “¿qué estados del receptor favorece, cuánto tiempo permanecen ocupados esos estados y qué proteínas están disponibles para acoplarse después?”. Una neurona rica en CB1 y una célula microglial que expresa CB2 no presentan el mismo entorno de señalización, incluso antes de considerar las diferencias en densidad de receptor.
Esta visión dinámica también debilita la antigua suposición de que los ligandos endógenos definen una sola línea de base fisiológica frente a la cual THC, cannabidiol o los cannabinoid sintéticos pueden clasificarse. THC es un fitocannabinoid con su propio patrón de eficacia y comportamiento cinético. Los endocannabinoids son señales impulsadas por eventos, a menudo fásicas y locales. Un agonista sintético de CB2 puede parecer selectivo en un ensayo recombinante, pero producir resultados inesperados en tejido si el receptor de reserva, el colesterol de membrana, la formación de heterómeros o el manejo por beta-arrestin difieren. La selectividad sobre el papel no es un destino.
Cómo la síntesis y la degradación locales modelan la interacción con el receptor
In vivo, la señalización de los endocannabinoids está gobernada tanto por la enzimología como por la farmacología de los receptores. En muchas sinapsis centrales, el 2-AG se produce postsinápticamente en respuesta a aumentos de calcio o a la activación de receptores acoplados a Gq/11, luego viaja de forma retrógrada hasta los receptores presinápticos CB1 y suprime la liberación de neurotransmisores. Se trata de un mecanismo de temporización medido en segundos, no de un tono difuso de fondo. La señal termina cuando MAGL y las hidrolasas relacionadas eliminan el 2-AG. La anandamida suele comportarse de forma distinta, con menor abundancia tisular, condiciones de liberación diferentes y una sensibilidad mayor a la degradación controlada por FAAH.
Eso significa que modificar una enzima puede alterar la interacción con el receptor sin cambiar el receptor en absoluto. La inhibición de FAAH eleva el tono de la anandamida, pero el resultado fisiológico depende de dónde se produzca la anandamida, de si TRPV1 también está presente y de si los receptores CB1 en ese microdominio están desensibilizados o internalizados. La inhibición de MAGL puede elevar de forma marcada el 2-AG, pero una elevación crónica puede inducir desensibilización de CB1 en algunos sistemas. Más ligando no siempre significa una señalización más útil.
El estudio de sistemas biológicos indexado como PMID: 42129940 refuerza este panorama más amplio al situar CB1 y CB2 entre nodos influyentes conectados con vías metabólicas, y no como interruptores aislados. Esa es la escala adecuada para comprender la acción de los endocannabinoids. La disponibilidad del ligando, el acceso a la membrana, la conformación del receptor, el acoplamiento a proteínas G, el reclutamiento de beta-arrestin y la degradación interactúan entre sí. También lo hacen los gradientes tisulares. CB2 no está simplemente “fuera del cerebro”, y CB1 no es solo “el receptor de la psicoactividad”. La señalización de los endocannabinoids es local, condicional y cinética. Cualquier explicación de la selectividad de subtipo que ignore esas características interpretará mal tanto la fisiología como el desarrollo de fármacos.
El sistema endocannabinoide como una red, no como un diagrama de dos receptores
El sistema endocannabinoide tiene poco sentido si se dibuja como dos círculos etiquetados CB1 y CB2 con flechas apuntando hacia afuera. Ese esquema ayudó al campo en sus inicios, especialmente después del trabajo de farmacología de receptores de Allyn Howlett y de la identificación de anandamide por Raphael Mechoulam y Lumír Hanuš, pero ahora oculta más de lo que revela. Las implicaciones prácticas son considerables. La World Health Organization estimó que 200 millones de personas usaron cannabis en 2019, aproximadamente el 4% de la población mundial de 15 a 64 años, mientras que la U.S. FDA afirma que, a fecha de 2025, hay un producto farmacológico derivado de cannabis y tres productos farmacológicos relacionados con cannabis aprobados. Un sistema de receptores que interviene en tantas exposiciones y tantas afirmaciones terapéuticas no puede tratarse como una simple pareja formada por un receptor cerebral y un receptor inmunitario.
Una visión de sistemas parte de un hecho básico: los endocannabinoids son lípidos producidos a demanda, degradados rápidamente e integrados en el mismo espacio bioquímico que los mediadores inflamatorios, las vías de remodelación de membranas y los circuitos de retroalimentación sináptica. Anandamide y 2-arachidonoylglycerol no son “mensajes” aislados. Se producen a partir de precursores de membrana, compiten con otras vías lipídicas por sustratos y actúan en tejidos donde la densidad de receptores, el tipo celular, la expresión enzimática y los socios de señalización difieren. Por eso el mismo ligando puede parecer ansiolítico en un contexto, intoxicante en otro, antiinflamatorio en otro y clínicamente decepcionante en un ensayo.
Qué aporta el análisis de red integrador
El estudio de análisis de red integrador indexado en PubMed y publicado en 2025/2026 (PMID: 42129940) es útil porque desplaza la atención desde los objetivos individuales hacia la organización del sistema. En lugar de preguntar solo qué ligando se une a CB1 o CB2, los autores mapean el sistema endocannabinoid como una red de interacciones que incluye receptores, enzimas biosintéticas y degradativas, intermediarios lipídicos y vínculos a nivel de vía con la inflamación, el metabolismo y la señalización neuronal. En ese marco, la influencia no se define solo por la abundancia de receptores. Se define por la conectividad y por la forma en que las perturbaciones se propagan a través de la red.
Eso importa para la interpretación. Si un fármaco modifica la señalización de CB1, el efecto descendente no se limita a la inhibición de la adenilil ciclasa mediada por Gi/o. Puede alterar la liberación de neurotransmisores, modificar el comportamiento de los canales de calcio y potasio, cambiar el reclutamiento de beta-arrestina, desencadenar la internalización del receptor y, de forma indirecta, modificar el flujo de sustratos lipídicos que también alimentan eicosanoids y otras vías inflamatorias. Un modelo de red captura mejor estas consecuencias entre sistemas que un esquema de ocupación de receptores.
Aquí también es donde se rompe el viejo atajo de “CB1 en el cerebro, CB2 en las células inmunitarias”. La distribución es graduada, específica de cada célula y dependiente del estado. CB1 es abundante en muchas poblaciones neuronales, pero no de forma uniforme en todo el cerebro, y también aparece en tejidos periféricos relevantes para el equilibrio energético, la nocicepción y la función de los órganos. CB2 está enriquecido en linajes inmunitarios, pero los trabajos más recientes sobre el sistema nervioso central han ido mucho más allá de la idea de que sea irrelevante para la biología cerebral. Una revisión de 2026 en Frontiers in Behavioral Neuroscience describió esto como “an update over the last 3 years”, subrayando la señalización de CB2 en contextos neuroinflamatorios y neurodegenerativos. La cuestión no es que CB2 se haya convertido de repente en “el receptor cerebral también”. La cuestión es que el significado del receptor depende de qué células estén activas, lesionadas, inflamadas o adaptándose.
El análisis de red ayuda a explicar por qué los fármacos dirigidos a receptores suelen producir efectos más amplios de lo esperado. Un fitocannabinoid activo en CB1 como delta-9-THC, un endocannabinoid como anandamide y un agonista sintético pueden activar el mismo receptor y, aun así, generar resultados temporales y espaciales distintos. La clase de ligando importa. También importan la eficacia, el tiempo de residencia, el metabolismo y el acceso a distintas conformaciones del receptor.
CB1 y CB2 como nodos influyentes
El estudio de red integrador identifica a CB1 y CB2 como nodos de gran influencia, no porque operen por separado, sino porque se sitúan en puntos de control donde convergen muchas vías. En términos de red, los nodos influyentes son lugares donde cambios locales pueden propagarse ampliamente. En términos de farmacología, son receptores cuyo estado de activación puede redirigir al mismo tiempo la señalización sináptica, las respuestas inmunitarias y los programas de estrés celular.
CB1 es el ejemplo más claro de por qué “influyente” no significa “simple”. Señaliza principalmente a través de proteínas Gi/o, pero eso solo es el comienzo. Según el ligando y el contexto, CB1 puede reclutar beta-arrestins, sufrir desensibilización dependiente de fosforilación, internalizarse, reciclarse o permanecer activamente en la membrana durante más tiempo. El artículo del American Journal of Psychiatry de 2025 sostiene que la señalización sesgada en CB1 es una estrategia terapéutica plausible para la esquizofrenia, un trastorno que la WHO dice que afecta a unos 24 millones de personas en todo el mundo. Esa propuesta solo tiene sentido si CB1 se entiende como un centro de señalización con salidas separables, no como un interruptor de encendido/apagado para la intoxicación. Un ligando que favorece una rama de señalización sobre otra puede preservar algunos efectos terapéuticos y reducir otros adversos. Puede, no necesariamente va a. El sesgo puede reducir problemas, pero no elimina el hecho de que CB1 forma parte de circuitos que gobiernan la cognición, la recompensa, el control motor, el procesamiento del dolor y el apetito.
CB2 muestra un problema similar desde la dirección opuesta. Durante mucho tiempo se lo presentó como el objetivo más seguro debido a su menor asociación con efectos psicoactivos agudos. Ese planteamiento es demasiado optimista. Los ligandos dirigidos a CB2 todavía pueden producir resultados decepcionantes o mixtos si la señalización relevante para la enfermedad depende del estado celular, la inducibilidad del receptor, el tono inflamatorio o la comunicación cruzada con enzimas metabólicas. La revisión estructural reciente en Frontiers in Chemical Biology, publicada en 2026, subraya que la selectividad entre “CB1 y CB2” depende de diferencias estructurales a nivel de receptor que modifican la unión, la eficacia y la regulación del receptor. El estudio indexado en PubMed de 2025/2026 sobre selectividad de subtipo (PMID: 41962866) va más allá al argumentar que la selectividad es dinámica y está determinada por el comportamiento conformacional, no por un encaje estático tipo llave-cerradura. Esa es una buena razón para dejar de hablar de compuestos selectivos para CB1 o para CB2 como si la selectividad fuera una propiedad fija con consecuencias biológicas fijas.
Vínculos entre la señalización de receptores y las vías metabólicas
La contribución más valiosa de la perspectiva de red es que reconecta la farmacología de receptores con el metabolismo lipídico. La señalización endocannabinoid se construye sobre una química relacionada con el ácido araquidónico. Anandamide y 2-AG se generan y degradan mediante sistemas enzimáticos que también influyen en la disponibilidad de lípidos bioactivos implicados en la inflamación y la respuesta tisular. Un fármaco que inhibe FAAH o MAGL no simplemente “eleva los endocannabinoids”. Redistribuye el tráfico de señalización a través de una red metabólica.
Eso puede ser útil. También puede salir mal. Elevar 2-AG, por ejemplo, puede potenciar la señalización de CB1 o CB2 en algunos compartimentos mientras altera las vías relacionadas con prostaglandins en otros, mediante desvío de sustratos u oxidación descendente. La implicación clínica es directa: las estrategias dirigidas a receptores y las dirigidas a enzimas pueden tener consecuencias a escala sistémica incluso cuando su objetivo de diseño es estrecho. Esta es una de las razones por las que los compuestos selectivos no se han traducido automáticamente en perfiles clínicos limpios en dolor, enfermedad psiquiátrica o trastornos inflamatorios, a pesar de la alta necesidad no cubierta. Solo el dolor crónico afecta a casi 1 de cada 5 adultos en Estados Unidos, según las National Academies, y aun así la farmacología de cannabinoid no ha producido una solución universalmente fiable basada en receptores.
Visto así, CB1 y CB2 se entienden mejor como nodos de control incrustados en circuitos lipídicos, inmunitarios y sinápticos. Su capacidad de ser objetivos farmacológicos es real. También lo es su capacidad de sorprender.
CB1 como diana farmacológica: promesa, riesgos y el problema de los efectos secundarios centrales
CB1 sigue siendo una de las dianas más tentadoras en la neurofarmacología porque se sitúa en un punto de control estratégico de la señalización sináptica. El trabajo de farmacología de receptores de Allyn Howlett estableció que los efectos de los cannabinoid no eran un fenómeno inespecífico de la membrana, sino una biología mediada por receptores, y el descubrimiento posterior de anandamida por Raphael Mechoulam y Lumír Hanuš proporcionó a ese receptor un sistema de señalización endógeno. Esa historia importa. Un receptor que ayuda a modular la liberación de neurotransmisores en circuitos del dolor, circuitos del apetito, vías del estrés y redes de recompensa parece un punto de apoyo plausible para varios trastornos complejos a la vez. También crea una trampa: cuando un receptor está integrado en tantas funciones cerebrales fundamentales, una manipulación amplia rara vez permanece confinada al efecto deseado.
Ese problema no es académico. La Organización Mundial de la Salud estima que 200 millones de personas usaron cannabis en 2019, aproximadamente el 4% de la población mundial de 15 a 64 años, por lo que la farmacología de CB1 ya opera a escala poblacional a través de cannabinoid vegetales, especialmente delta-9-THC. Sin embargo, los medicamentos cannabinoid aprobados siguen siendo pocos: la FDA de EE. UU. indica que, a fecha de 2025, ha aprobado un producto farmacéutico derivado de cannabis y tres productos farmacéuticos relacionados con cannabis. La brecha entre la exposición generalizada y la escasez de terapias aprobadas dice algo importante sobre CB1. La biología es convincente. La terapéutica es difícil.
Fundamentación terapéutica en dolor, apetito y neuropsiquiatría
La atracción comienza por dónde actúa CB1 y qué hace. CB1 se expresa en alta densidad en muchas terminales presinápticas del sistema nervioso central, donde los endocannabinoid suelen producirse a demanda en células postsinápticas y viajar en sentido retrógrado a través de la sinapsis para suprimir la liberación de neurotransmisores. Este sistema retrógrado puede atenuar glutamato, GABA y otras vías de señalización según el tipo celular y el circuito. Para el dolor, esa es una fundamentación obvia: si casi 1 de cada 5 adultos en Estados Unidos vive con dolor crónico, como señaló las Academias Nacionales en 2017, un receptor capaz de reducir la transmisión nociceptiva y modificar el componente afectivo del dolor inevitablemente atraerá atención.
Pero CB1 no es un simple interruptor analgésico. En algunos circuitos, la supresión de la liberación inhibitoria de GABA puede desinhibir neuronas postsinápticas en lugar de silenciarlas. En otros, predomina la supresión glutamatérgica. El efecto depende de la sinapsis, del estado de la red y del ligando. Los endocannabinoid como anandamida y 2-AG son señales breves, generadas localmente. Los phytocannabinoid y los agonistas sintéticos llegan con cinéticas distintas, distinta eficacia y, a menudo, un compromiso mucho más amplio con receptores a lo largo del tiempo. Esa diferencia ayuda a explicar por qué “la activación de CB1 reduce el dolor” es demasiado simplista para guiar un diseño clínico seguro.
El apetito es una segunda gran fundamentación. La señalización de CB1 promueve la ingesta de alimentos e interactúa con circuitos hipotalámicos y mesolímbicos que vinculan la homeostasis energética con la recompensa. Esto convirtió el antagonismo de CB1 en una estrategia antiobesidad atractiva y la agonismo de CB1, o su potenciación indirecta, en una vía potencial para la caquexia o la pérdida de apetito. La lógica no era irracional. Se apoyaba en la fisiología básica. Sin embargo, el apetito es inseparable del estado de ánimo, la saliencia y el procesamiento del estrés, todos ellos también modulados por CB1.
La neuropsiquiatría es donde el receptor se vuelve más interesante y más peligroso. CB1 modula circuitos relevantes para la dopamina, la inhibición cortical, la plasticidad hipocampal, el aprendizaje del miedo y la respuesta al estrés. Eso le confiere relevancia teórica para los trastornos de ansiedad, las condiciones relacionadas con trauma, la depresión, los trastornos por consumo de sustancias y la enfermedad psicótica. El American Journal of Psychiatry sostuvo en 2025 que la señalización sesgada de CB1 es una estrategia plausible para la esquizofrenia, un trastorno que afecta a unas 24 millones de personas en todo el mundo según la WHO. La importancia de ese artículo no es que un fármaco de CB1 para la esquizofrenia esté listo. Es que el campo ha avanzado desde la agonismo y antagonismo crudos hacia una farmacología selectiva por vías, adoptando el enfoque más amplio de los GPCR.
Por qué la diana CB1 es científicamente atractiva pero clínicamente difícil
CB1 es científicamente atractivo precisamente por la misma razón por la que es clínicamente difícil: es un nodo de señalización denso. A nivel canónico, CB1 se acopla principalmente a proteínas Gi/o, reduciendo la actividad de la adenilato ciclasa, modulando canales iónicos y suprimiendo la liberación de neurotransmisores. Eso por sí solo bastaría para hacerlo importante. Pero además recluta beta-arrestinas, experimenta desensibilización e internalización, y puede ser moldeado por moduladores alostéricos y por estados conformacionales específicos del ligando. La revisión estructural de Frontiers in Chemical Biology de 2026 lo expone con claridad para CB1 y CB2: la selectividad y la eficacia de los ligandos surgen de diferencias estructurales a nivel del receptor que afectan la unión, la señalización y la regulación del receptor. Por tanto, cannabinoid de apariencia similar pueden producir farmacología significativamente distinta.
Eso no es solo un detalle de química medicinal. Va directamente a los efectos secundarios. Un ligando que impulsa con fuerza una conformación activa puede producir más internalización del receptor, más tolerancia o un perfil conductual distinto al de un ligando que estabiliza otro estado. Un estudio indexado en PubMed de 2025/2026 sobre selectividad por subtipos argumentó además que la selectividad de los endocannabinoid es dinámica, no un evento fijo de llave y cerradura, reforzando la idea de que el comportamiento del receptor cambia con el contexto conformacional. Los desarrolladores de fármacos no se enfrentan a una única diana CB1. Se enfrentan a un conjunto móvil.
Por eso los agonistas y antagonistas inespecíficos de CB1 han decepcionado con tanta frecuencia. Una agonismo central intenso puede deteriorar la memoria, la atención, el rendimiento psicomotor y la regulación de la ansiedad, además de activar vías de recompensa de formas que complican el riesgo de dependencia. Un antagonismo central intenso puede reducir el apetito, sí, pero también puede interferir con los mismos circuitos de amortiguación del estrés y del afecto que los cannabinoid endógenos estabilizan normalmente. CB1 participa profundamente en la cognición, la recompensa y la regulación del estrés. Cualquier fármaco que empuje ese sistema con demasiada fuerza en una sola dirección debe considerarse arriesgado hasta demostrar lo contrario.
La biología de sistemas respalda esta cautela. Un análisis de redes integrativo de 2025/2026 identificó CB1 y CB2 como nodos muy influyentes en el sistema endocannabinoid y conectó la señalización del receptor con vías metabólicas más amplias. Ese hallazgo va en contra de la simplificación centrada en un solo receptor. Alterar CB1 no solo desplaza una sinapsis. Puede propagarse a través de redes endocrinas, inmunitarias, metabólicas y conductuales.
Lecciones de las estrategias de acción central frente a las restringidas a la periferia
La lección más clara de la primera era del desarrollo de fármacos dirigidos a CB1 es que la exposición central suele ser el punto donde la eficacia y la toxicidad se vuelven inseparables. Los agonistas de CB1 con acción central pueden intervenir en circuitos del dolor y del apetito, pero la misma penetración cerebral eleva la probabilidad de efectos tipo intoxicación, sedación, alteración cognitiva y eventos adversos psiquiátricos. Los antagonistas centrales de CB1 ilustran la falla opuesta: pueden alcanzar objetivos metabólicos mientras producen efectos inaceptables sobre el estado de ánimo y la ansiedad, porque también bloquean el tono endocannabinoid endógeno en circuitos límbicos.
Esa historia explica por qué los ligandos de CB1 restringidos a la periferia se volvieron atractivos. La idea es sencilla: si algunos beneficios terapéuticos dependen de CB1 en tejidos periféricos como las aferencias primarias, el intestino, el hígado, el tejido adiposo u otros sitios fuera del SNC, entonces mantener un fármaco fuera del cerebro podría preservar los efectos útiles y reducir los efectos adversos centrales. Para el dolor, esto podría significar actuar sobre mecanismos nociceptivos periféricos. Para la enfermedad metabólica, podría implicar reducir la lipogénesis o modificar vías relacionadas con la glucosa sin alterar el estado de ánimo ni la cognición.
La lógica es sólida, pero no es una solución mágica. Primero, la biología de CB1 periférica y central no está separada de manera limpia en estados patológicos. El dolor crónico, la alimentación y el estrés implican bucles entre cerebro y cuerpo. Segundo, la exclusión por la barrera hematoencefálica es un problema cuantitativo, no binario. Incluso pequeñas cantidades de penetración en el SNC pueden importar, especialmente con dosificación crónica. Tercero, un ligando de CB1 verdaderamente periférico aún puede fracasar clínicamente si la enfermedad diana depende en gran medida de circuitos centrales o si las vías compensatorias atenúan la eficacia.
La estrategia más reciente no es simplemente “mantenerse fuera del cerebro”. Es “dar forma a la señal”. Eso incluye agonistas parciales, antagonistas neutrales en lugar de agonistas inversos, moduladores alostéricos y ligandos sesgados que favorecen determinadas vías descendentes sobre otras. La discusión de 2025 en el American Journal of Psychiatry sobre la señalización sesgada de CB1 en la esquizofrenia encaja aquí. Si un ligando puede preservar señales terapéuticas seleccionadas mientras reduce el reclutamiento de beta-arrestinas, la desensibilización del receptor o los efectos indeseados sobre los circuitos, entonces CB1 podría volverse más manejable. Podría. Sigue sin haber garantía de que el sesgo de vía observado en sistemas celulares se traduzca con precisión a ventanas terapéuticas humanas.
Por tanto, la visión actual de CB1 como diana farmacológica no es ni despectiva ni romántica. CB1 resulta atractivo porque regula la ganancia sináptica precisamente en los tipos de circuitos que los clínicos desean modular. Es difícil porque esos mismos circuitos también gobiernan la memoria, el afecto, la motivación y la resiliencia al estrés. Los ligandos restringidos a la periferia, las estrategias de señalización selectiva y una mejor comprensión estructural han mejorado la lógica de diseño. No han eliminado el problema de fondo. Un receptor tan central para la función cerebral rara vez tolera una farmacología burda.
CB2 como diana terapéutica: neuroinflamación, neurodegeneración y modulación inmunitaria
CB2 se ha convertido en una de las dianas más tentadoras de la farmacología del cannabinoid por una razón simple: parece ofrecer acceso a la biología inmunitaria y glial con menor riesgo de intoxicación, deterioro de la memoria y potencial de abuso clásicamente asociados con una fuerte activación de CB1. Ese atractivo importa mucho más allá del mapeo académico de receptores. La Organización Mundial de la Salud estimó que 200 millones de personas consumieron cannabis en 2019, es decir, el 4% de la población mundial de 15 a 64 años, y la FDA de EE. UU. todavía enumera solo un producto farmacéutico derivado de cannabis y tres productos farmacéuticos relacionados con cannabis como aprobados a fecha de 2025. El interés es alto. La traslación es difícil.
La antigua simplificación decía que CB1 es el receptor cerebral y CB2 es el receptor inmunitario. Ahora eso es demasiado burdo para resultar útil. La revisión de 2026 en Frontiers in Behavioral Neuroscience, formulada explícitamente como “una actualización de los últimos 3 años”, considera CB2 relevante para los trastornos del sistema nervioso central precisamente porque participa en mecanismos neuroinflamatorios y neurodegenerativos, incluidas la activación microglial, la señalización de citocinas y la comunicación cruzada neurona-glía. El cambio clave no es que CB2 haya pasado de repente a ser abundante en todo el cerebro. No lo hizo. El cambio es que una baja expresión basal en muchas regiones del SNC no implica irrelevancia funcional, especialmente cuando los estados patológicos pueden alterar la expresión del receptor, su tráfico y su señalización.
Por qué CB2 suele considerarse una diana más segura
El argumento de seguridad para CB2 comienza con la distribución, pero no termina ahí. CB2 está enriquecido en células inmunitarias y suele estar sobreexpresado en microglía activada, macrófagos infiltrantes y otras poblaciones celulares inflamatorias. En contraste, CB1 se expresa densamente en terminales presinápticos de la corteza, el hipocampo, los ganglios basales y el cerebelo, donde moldea de forma intensa la liberación de neurotransmisores. Esa diferencia anatómica importa porque un ligando que interactúe principalmente con CB2 tiene menos probabilidades de reproducir el perfil psicoactivo clásico asociado con CB1.
Menos probabilidades no es lo mismo que imposibilidad. La selectividad es un objetivo móvil. La revisión de 2026 en Frontiers in Chemical Biology subraya que la selectividad entre “CB1 y CB2” depende de diferencias estructurales que alteran la unión, la eficacia y la regulación del receptor, y un estudio reciente indexado en PubMed sobre selectividad de subtipos sostiene que la discriminación de endocannabinoid entre subtipos de receptor refleja dinámicas conformacionales más que un modelo fijo de llave-cerradura. En la práctica, eso significa que un compuesto puede parecer selectivo para CB2 en un ensayo y comportarse de forma menos limpia en otro, especialmente cuando la densidad del receptor, el entorno de membrana y los transductores aguas abajo difieren. El mismo ligando puede sesgar la señalización de proteínas G en un tipo celular, reclutar beta-arrestinas en otro y mostrar distintas cinéticas de desensibilización o internalización en condiciones inflamatorias.
Esa complejidad mecanística, de hecho, refuerza el caso de perseguir CB2 con cautela. Si el objetivo terapéutico es reducir la salida de citocinas de la microglía, limitar el reclutamiento de células inmunitarias periféricas o alterar el estrés metabólico glial sin perturbar de forma intensa la transmisión sináptica cortical, entonces CB2 sigue siendo el receptor inicial más plausible al que dirigir el tratamiento. La señalización sesgada de CB1, como se argumenta en el artículo de 2025 del American Journal of Psychiatry sobre esquizofrenia, puede ampliar con el tiempo la ventana terapéutica de los fármacos dirigidos a CB1. Pero ese es un argumento a favor de una farmacología más inteligente de CB1, no una razón para ignorar la menor carga de psicoactividad de los enfoques centrados en CB2.
Sin embargo, más seguro no debe confundirse con más eficaz. Un fármaco puede evitar una intoxicación evidente y aun así fracasar porque el receptor se expresa de manera demasiado escasa, demasiado variable o demasiado transitoria durante el proceso patológico como para generar un efecto clínico significativo. Eso ha ocurrido repetidamente en el desarrollo de fármacos contra la neuroinflamación, no solo en la ciencia cannabinoid.
Evidencia y límites en modelos de enfermedad del SNC
La revisión de Frontiers in Behavioral Neuroscience expone por qué CB2 sigue reapareciendo en modelos de enfermedad de Alzheimer, enfermedad de Parkinson, esclerosis múltiple, traumatismo craneoencefálico, ictus y dolor neuropático: todas estas afecciones incluyen señalización inflamatoria, reactividad glial, estrés oxidativo y daño tisular progresivo. En muchos estudios con roedores, los agonistas de CB2 reducen mediadores proinflamatorios como TNF-α, IL-1β e IL-6, desplazan la microglía hacia fenotipos menos dañinos o atenúan la infiltración leucocitaria. A veces eso se acompaña de una mejor conducta motora, una reducción del tamaño de la lesión o la preservación de marcadores sinápticos. Son señales reales, no artefactos que deban descartarse.
Pero el balance es desigual. Los efectos a menudo dependen del momento de administración. Un agonista de CB2 administrado durante la fase inflamatoria temprana tras una lesión puede hacer algo muy distinto del mismo agonista administrado durante la degeneración crónica. La dosis también importa, al igual que la clase de ligando. Los endocannabinoid como 2-AG y anandamida no son intercambiables con los fitocannabinoid ni con agonistas sintéticos con preferencia por CB2. Sus concentraciones locales, su degradación metabólica y sus acciones fuera del objetivo también difieren. Lo mismo ocurre con sus preferencias de vía.
Los propios modelos de enfermedad son otra limitación. Un ratón transgénico de Alzheimer con patología amiloide y microglía reactiva no es la enfermedad de Alzheimer en un ser humano de 74 años con fracaso mixto vascular, inflamatorio y de proteostasis. La encefalomielitis autoinmune experimental es útil para la biología de la esclerosis múltiple, pero captura solo una parte de la condición humana. Por tanto, los resultados preclínicos positivos establecen plausibilidad, no prueba.
También persiste un problema de medición en la biología de CB2. La especificidad de los anticuerpos ha sido durante mucho tiempo un punto débil, lo que dificulta confiar en las afirmaciones sobre localización del receptor más de lo que muchos artículos sugieren. Los bajos niveles de expresión en el cerebro sano, la expresión inducible en la enfermedad y las diferencias entre especies complican aún más la interpretación. Algunos hallazgos de “CB2 neuronal” informados pueden ser reales en contextos específicos, pero las afirmaciones amplias sobre una expresión neuronal constitutiva extendida de CB2 siguen siendo menos sólidas de lo que el campo sugirió en su momento.
Aquí es donde la señalización importa tanto como la anatomía. CB2 es un GPCR acoplado a Gi/o, pero ese resumen oculta variaciones importantes: la inhibición de la adenylyl cyclase, la modulación de las vías MAPK, los efectos sobre los canales iónicos, el reclutamiento de beta-arrestina, la desensibilización del receptor y la internalización pueden alterar el resultado biológico neto. Un ligando que reduce la liberación de citocinas en microglía cultivada puede fracasar in vivo si desensibiliza rápidamente los receptores o si desencadena un programa dominado por arrestina que resulte poco útil. Un menor riesgo de intoxicación no resuelve estos problemas farmacológicos.
Lo que sugiere la literatura reciente para las terapias futuras
La literatura reciente se aleja de la antigua búsqueda de un único “agonista de CB2 para la inflamación cerebral” y se orienta hacia una estrategia más selectiva y dependiente del contexto. La revisión estructural de 2026 y el artículo de 2025/2026 sobre selectividad de subtipos respaldan la misma lección: la diana del receptor dependerá del estado conformacional, el perfil de eficacia y el contexto tisular, no solo de una afinidad nominal por el subtipo. El diseño de fármacos avanza hacia ligandos selectivos en función, no solo en unión.
Eso probablemente signifique tres cosas. Primero, los futuros fármacos de CB2 necesitarán mejores huellas de señalización, incluyendo el sesgo entre proteínas G y beta-arrestina, el tiempo de residencia en el receptor y los efectos específicos de tipo celular en sistemas humanos, no solo en líneas de ensayo inmortalizadas. Segundo, la estratificación de la enfermedad será importante. Una terapia dirigida a CB2 puede funcionar mejor allí donde la amplificación inflamatoria sea un impulsor principal, y no solo un subproducto, de la patología. Tercero, pensar en combinaciones es inevitable. Un análisis integrador de redes publicado en un estudio reciente indexado en PubMed identificó CB1 y CB2 como nodos influyentes en vías más amplias de endocannabinoid y metabólicas, lo que encaja con la realidad vivida de las enfermedades del SNC: inflamación, estrés mitocondrial, señalización lipídica y disfunción sináptica están entrelazadas.
En condiciones inflamatorias específicas, la modulación de CB2 puede reducir la señalización inmunitaria dañina mientras evita gran parte de la carga psicoactiva vinculada con CB1.Preliminary evidence
Eso no convierte a CB2 en un callejón sin salida. Lo convierte en una diana que probablemente pertenece a la farmacología de precisión más que a la mitología de los receptores. El caso actual más sólido no es “la activación de CB2 trata la neurodegeneración”. Es más estrecho: bajo condiciones inflamatorias específicas, en poblaciones celulares específicas, y con las propiedades adecuadas del ligando, la modulación de CB2 puede reducir la señalización inmunitaria dañina al tiempo que evita buena parte de la carga psicoactiva vinculada a CB1. Esa es una hipótesis terapéutica creíble. Todavía no es una regla clínicamente validada.
Compuestos del cannabis en CB1 y CB2: THC, CBD, cannabinoides menores y ligandos sintéticos
cannabis no actúa a través de una sola sustancia química ni de un único mecanismo receptor. Esa afirmación parece obvia, y aun así el debate público sigue reduciendo la farmacología a un emparejamiento falso: THC equivale a CB1, CBD equivale a CB2. La literatura no respalda esa simplificación. cannabis contiene muchos fitocannabinoides, además de terpenes, flavonoides y otros constituyentes, mientras que el cuerpo produce sus propios endocannabinoids, como la anandamida y el 2-arachidonoylglycerol, identificados mediante trabajos asociados con Raphael Mechoulam, Lumír Hanuš y otros. Además, la mayor parte de lo que los científicos saben sobre activación receptorial, eficacia, selectividad, desensibilización y sesgo de señalización procede de ligandos sintéticos diseñados como sondas o candidatos a fármacos, más que de los compuestos vegetales por sí solos.
Esa distinción importa porque CB1 y CB2 no son cerraduras pasivas que esperan una llave cannabinoid. Son GPCR de clase A con múltiples estados conformacionales, preferencias de acoplamiento y destinos regulatorios. Una revisión estructural de 2026 en Frontiers in Chemical Biology lo señala con claridad: la selectividad de los ligandos en CB1 y CB2 depende de diferencias estructurales a nivel del receptor que modifican la unión, la eficacia y la regulación del receptor, no solo de si una molécula “actúa sobre” uno u otro receptor. Un estudio reciente indexado en PubMed sobre selectividad de subtipos sostiene asimismo que la preferencia de los endocannabinoids por CB1 frente a CB2 surge de la dinámica conformacional, y no de un simple ajuste de llave y cerradura. Por tanto, cuando dos cannabinoid parecen químicamente similares pero producen efectos distintos en el cerebro, el tejido inmunitario o los circuitos del dolor, eso no es un detalle secundario. Es la farmacología central.
Dado que la World Health Organization estimó que 200 millones de personas usaron cannabis en 2019, aproximadamente el 4 % de la población mundial de 15 a 64 años, estas distinciones no son trivialidades académicas. Modelan la intoxicación, las afirmaciones terapéuticas, la seguridad y el motivo por el cual algunos fármacos dirigidos a receptores tienen éxito mientras otros fracasan.
Cómo THC interactúa con CB1 y contribuye a los efectos psicoactivos
Delta-9-tetrahydrocannabinol, o THC, sigue siendo el compuesto central para entender la intoxicación por cannabis porque interactúa directamente con CB1 en el sistema nervioso central. Esa es la explicación más clara a nivel receptor de los efectos psicoactivos característicos del cannabis, aunque “psicoactivo” engloba varios resultados separables: alteración de la percepción del tiempo, recompensa, alteración de la memoria, ansiedad en algunos usuarios e impedimento de la coordinación motora. El trabajo farmacológico sobre receptores de Allyn Howlett ayudó a establecer este marco centrado en CB1, y sigue siendo fundamental.
Pero “THC activa CB1” es solo el comienzo. THC no es un interruptor perfecto de encendido. Por lo general se describe como un agonista parcial en CB1, lo que significa que su eficacia depende de la densidad del receptor, del contexto tisular y del ensayo de señalización utilizado. En regiones con alta expresión de CB1, como la corteza, el hipocampo, los ganglios basales y el cerebelo, eso puede bastar para producir efectos centrales intensos. A nivel sináptico, CB1 suele ser presináptico, donde su activación suprime la liberación de neurotransmisores. Eso significa que THC puede amortiguar la liberación de glutamato en un circuito, la liberación de GABA en otro, y producir resultados de red opuestos dependiendo de qué células expresan el receptor y en qué momento.
Por eso la localización del receptor no puede reducirse a “CB1 en el cerebro”. CB1 es abundante en el cerebro, sí, pero no de forma uniforme, no es estático y no es funcionalmente idéntico entre tipos neuronales. El resultado es que los efectos de THC dependen del circuito. La euforia y el refuerzo forman parte del cuadro. También lo hacen la alteración de la memoria a corto plazo y la señalización relevante para psicosis en personas susceptibles.
Ese último punto ha adquirido más importancia, no menos. Un artículo de 2025 en American Journal of Psychiatry argumentó que la señalización sesgada de CB1 es una estrategia terapéutica plausible en la esquizofrenia, precisamente porque la biología de CB1 no puede entenderse como una activación receptorial unidimensional. La esquizofrenia afecta a unos 24 millones de personas en todo el mundo según la WHO, y cualquier discusión seria sobre THC en CB1 debe tener en cuenta que la misma familia de receptores implicada en la intoxicación también se estudia mediante agonismo sesgado, reclutamiento de beta-arrestin y diseño farmacológico selectivo de vías. THC puede activar la señalización acoplada a Gi/o, pero CB1 también sufre fosforilación, desensibilización e internalización, con consecuencias para la tolerancia y la respuesta posterior. La dosis y el momento importan. La exposición repetida cambia el sistema.
Los ligandos sintéticos de CB1 dejaron esto claro hace tiempo. Compuestos como CP55,940 y WIN55,212-2 son agonistas de investigación potentes que a menudo producen respuestas receptoras más intensas o más limpias que THC en ensayos de laboratorio. Rimonabant, un inverse agonist de CB1, mostró la dirección opuesta: bloquear o empujar CB1 por debajo de su actividad basal podía reducir el apetito y el peso corporal, pero los efectos adversos psiquiátricos hundieron el fármaco. Esa historia clínica es una advertencia. La selección de CB1 puede ser farmacológicamente elegante y, aun así, médicamente decepcionante.
Por qué CBD no encaja en una historia simple de agonista de CB1/CB2
CBD es el cannabinoid que con más frecuencia se simplifica hasta convertirse en un mito. Se presenta de forma rutinaria como “el no intoxicante que actúa a través de CB2” o como el opuesto directo de THC en cuanto a receptores. Ninguna de esas afirmaciones es exacta.
| Clase de compuesto o ejemplo | Cómo describe el artículo el comportamiento del receptor | Idea clave |
|---|---|---|
| THC | Agonista parcial de CB1 y CB2; central para la intoxicación a través de CB1 | La interacción con CB1 ayuda a explicar los efectos psicoactivos, pero el resultado sigue siendo dependiente del contexto |
| CBD | No es un agonista directo de CB1 o CB2; puede actuar de forma alostérica o indirecta | CBD no es un simple espejo de THC respecto de CB2 |
| Cannabinoides menores | Los efectos informados varían según el ensayo y la concentración | No generalizar el comportamiento del receptor de un cannabinoide a todos |
| Ligandos sintéticos | A menudo se usan para definir eficacia, selectividad y sesgo | Los compuestos sonda pueden comportarse de forma muy diferente a los cannabinoides vegetales |
CBD no se comporta como un agonista directo y simple de CB1 o CB2 de la manera en que THC lo hace en CB1. Su afinidad por los sitios ortostéricos de CB1 y CB2 es relativamente baja, y gran parte de su farmacología parece implicar acciones indirectas, alostéricas o sobre receptores distintos de cannabinoid. Según el sistema estudiado, se ha informado que CBD actúa como modulador alostérico negativo en CB1, lo que podría alterar cómo señalizan THC o los endocannabinoids sin limitarse a encender o apagar el receptor. Esa distinción es crucial. Modular la forma del receptor y la eficiencia de la señalización no es lo mismo que el agonismo clásico.
CBD también interactúa con dianas más allá de CB1 y CB2, incluidas TRPV1, 5-HT1A, PPAR-gamma, vías relacionadas con la adenosina y enzimas implicadas en el tono endocannabinoid. Algunos efectos pueden reflejar una señalización alterada de anandamida más que una estimulación directa del receptor. Eso ayuda a explicar por qué el perfil clínico de CBD no encaja de forma limpia con la antigua abreviatura basada en receptores. La U.S. FDA señala que ha aprobado un producto farmacológico derivado de cannabis y tres productos farmacológicos relacionados con cannabis a partir de 2025; el uso médico mejor establecido de CBD se encuentra en la epilepsia, una afección que afecta a alrededor de 50 millones de personas en todo el mundo, y esa vía terapéutica no puede explicarse con un modelo caricaturesco de “CBD se une a CB2 y reduce la inflamación”.
Incluso para CB2, el panorama es más amplio que la narrativa periférica más antigua. Una revisión de 2026 en Frontiers in Behavioral Neuroscience describió “una actualización de los últimos 3 años” en la que la señalización de CB2 ganó atención en trastornos del sistema nervioso central por sus vínculos con mecanismos neuroinflamatorios y neurodegenerativos. Eso no significa que CBD sea simplemente un fármaco de CB2. Significa que el propio receptor tiene una huella funcional mayor de la que sugiere la historia pública.
La importancia de separar los compuestos vegetales de los mitos receptoriales
Los phytocannabinoids no son intercambiables, y tampoco son intercambiables con los ligandos sintéticos. Se ha informado que cannabinoid menores como cannabigerol (CBG), cannabinol (CBN), tetrahydrocannabivarin (THCV) y cannabichromene (CBC) interactúan con CB1 y CB2 de formas que varían según el ensayo y la concentración. Algunos muestran agonismo débil, otros agonismo parcial, otros antagonismo en determinadas condiciones y otros una actividad relevante en dianas no cannabinoid. THCV es un buen ejemplo: en algunos sistemas se ha descrito como antagonista de CB1 o antagonista neutro a dosis bajas y como agonista parcial a dosis más altas. Eso no es inconsistencia por sí misma. Refleja una farmacología receptorial dependiente del contexto.
Las herramientas de investigación afinan este punto. Los endocannabinoids, los phytocannabinoids y los ligandos sintéticos pueden activar “CB1 y CB2”, pero no estabilizan las mismas conformaciones receptoras ni producen el mismo sesgo a través de proteínas G y beta-arrestins. La revisión estructural de 2026 y los análisis de red recientes indexados en PubMed empujan al campo lejos de la mitología receptorial y hacia la farmacología de sistemas. Los receptores se sitúan dentro de redes de señalización específicas de cada célula, vías metabólicas y maquinaria de tráfico. El perfil clínico de un ligando emerge de ese contexto completo, no de una etiqueta como “CB2 antiinflamatorio” o “CBD equilibra THC”.
Para una cannabis wiki, esa es la línea más importante: THC tiene una relevancia principal para la intoxicación mediada por CB1, pero la farmacología del cannabis no termina ahí, y CBD no es una simple imagen especular de THC en CB2. Los compuestos vegetales inician la historia. El estado del receptor, la distribución tisular, la eficacia, el sesgo y la regulación deciden cómo termina.
Esquizofrenia, riesgo de psicosis y lo que la biología de CB1 implica y no implica
Por qué la esquizofrenia entra en la conversación sobre receptores
La esquizofrenia es importante aquí porque CB1 no es solo otro receptor en una larga lista de dianas. Es uno de los receptores acoplados a proteína G más abundantes en el cerebro, situado en terminales presinápticas donde puede atenuar la liberación de neurotransmisores y remodelar la actividad de red en circuitos ya implicados en la psicosis: corteza, hipocampo, estriado, amígdala. Cuando un receptor se sitúa tan cerca del control de circuitos relacionados con glutamato, GABA, dopamina y la respuesta al estrés, los investigadores en psiquiatría prestan atención.
La escala de salud pública hace que esto sea más que una cuestión académica. La Organización Mundial de la Salud estima que cannabis fue utilizado por 200 millones de personas en 2019, o el 4% de la población mundial de 15 a 64 años. La WHO también estima que la esquizofrenia afecta a unos 24 millones de personas en todo el mundo, aproximadamente 1 de cada 300 personas. No se trata de poblaciones pequeñas y aisladas. Se solapan en clínicas reales, en servicios de urgencias y en estudios de cohortes longitudinales. Cualquier biología de receptores que pueda arrojar luz sobre el riesgo de psicosis, el desarrollo de fármacos antipsicóticos o ambos merece un examen serio.
Ese es el contexto traslacional del artículo de 2025 en American Journal of Psychiatry que sostiene que la señalización sesgada de CB1 es una estrategia terapéutica plausible para la esquizofrenia. La idea central es fácil de enunciar y fácil de malinterpretar. Fácil de enunciar: CB1 no se comporta como un simple interruptor de encendido y apagado, y distintos ligandos pueden estabilizar diferentes conformaciones del receptor, desviando la señalización hacia vías intracelulares específicas o alejándola de ellas. Fácil de malinterpretar: si CB1 es relevante para la investigación del tratamiento de la esquizofrenia, algunos lectores saltan a la afirmación falsa de que “la activación de cannabinoid ayuda a la esquizofrenia” o de que el consumo de cannabis es, por tanto, medicinal por defecto. El artículo no respalda ese salto.
Históricamente, la ciencia del cannabinoid quedó atrapada en abreviaturas simplificadoras. CB1 se presentó como el “receptor cerebral”, CB2 como el “receptor inmunitario”, y los efectos psicoactivos se atribuyeron casi por completo a la ocupación de CB1. Ese marco siempre fue incompleto y, en 2025, resulta claramente demasiado tosco. Los trabajos fundacionales de Allyn Howlett establecieron la farmacología de los receptores cannabinoid, mientras que Mechoulam y Hanus ayudaron a definir la biología de endocannabinoid a través de la anandamida. Desde entonces, la farmacología de receptores ha pasado de la presencia del receptor al estado del receptor. Ese cambio importa para la esquizofrenia porque la fisiopatología no está causada solo por la expresión del receptor. Surge del momento, el tipo celular, el contexto de circuito y la respuesta intracelular.
La literatura más reciente refuerza ese punto. Una revisión de 2026 en Frontiers in Chemical Biology explica que las diferencias estructurales entre CB1 y CB2 condicionan la unión de ligandos, la eficacia, la señalización y la regulación del receptor. Un estudio indexado en PubMed de 2025/2026 sobre la selectividad de subtipos sostiene que la selectividad de endocannabinoid puede surgir de la dinámica conformacional más que de un modelo estático de llave y cerradura. Otro trabajo de análisis de redes de 2025/2026 identifica a CB1 y CB2 como nodos de gran influencia en todo el sistema endocannabinoid, vinculados a vías metabólicas y de señalización más que a compartimentos receptoriales aislados. Para la investigación sobre esquizofrenia, esto significa que hay que dejar de preguntar solo “¿este compuesto actúa sobre CB1?” y empezar a preguntar “¿sobre qué estados de CB1, en qué células y bajo qué condiciones de exposición?”
De ahí proviene la seriedad clínica. No del lenguaje de receptor a nivel de eslogan. Sino de la precisión mecanística.
Señalización sesgada frente a activación brusca del receptor
La activación brusca de CB1 es un modelo deficiente para razonar terapéuticamente. Condensa demasiadas variables en una sola palabra: activación. Los ligandos endógenos como la anandamida y el 2-AG, los fitocannabinoids como delta-9-THC y los agonistas sintéticos no se comportan de manera idéntica en CB1, aunque todos cuenten como ligandos. Pueden diferir en afinidad, eficacia, tiempo de permanencia, exposición regional y sesgo de vía. También pueden diferir en la fuerza con la que reclutan proteínas Gi/o frente a beta-arrestinas, en la rapidez con la que desencadenan desensibilización y en el grado de internalización del receptor tras dosis repetidas.
Esa distinción es central para el argumento sobre esquizofrenia en AJP. Un ligando que desplaza suavemente a CB1 hacia un programa intracelular puede no parecerse a otro que impulsa una activación amplia del receptor en muchas regiones cerebrales. En farmacología de GPCR, esa diferencia puede separar un fármaco útil de uno intolerable. CB1 es especialmente sensible a este problema porque el mismo receptor puede reducir la transmisión excitadora en una sinapsis, suprimir la transmisión inhibidora en otra y alterar la plasticidad descendente de formas que no son funcionalmente equivalentes. Pequeñas diferencias moleculares pueden producir grandes consecuencias a nivel de sistema.
THC es el ejemplo obvio de por qué “diana CB1” no equivale a “terapia CB1”. THC es un fitocannabinoid con propiedades de agonista parcial en CB1, pero los efectos vividos y clínicos de THC no son una lectura pura de una sola rama de señalización. La dosis importa. La edad de exposición importa. La exposición repetida importa. También importa la proporción de THC respecto de otros componentes y si la persona tiene vulnerabilidad psiquiátrica. Los efectos agudos parecidos a intoxicación, la ansiedad, el cambio perceptivo y los síntomas psicóticos transitorios en algunos usuarios no nos dicen que todos los terapéuticos dirigidos a CB1 estén condenados. Sí nos dicen que la estimulación indiscriminada de CB1 no es un atajo hacia el tratamiento antipsicótico.
La agonía sesgada es atractiva precisamente porque intenta evitar esa crudeza. La esperanza terapéutica no es “más CB1”. Es “el perfil de señalización correcto de CB1”. Eso puede significar favorecer vías de señalización vinculadas a la estabilización de circuitos mientras se limitan las vías relacionadas con efectos cognitivos o psicotomiméticos adversos, tolerancia o desregulación del receptor. Si esto puede lograrse en pacientes sigue siendo una pregunta abierta. El concepto es plausible. No es un éxito clínico demostrado.
CB2 también pertenece a este panorama, aunque no como solución simplista. La revisión de 2026 en Frontiers in Behavioral Neuroscience describe “una actualización de los últimos 3 años” en la que la señalización de CB2 ha ganado atención en trastornos del sistema nervioso central por sus vínculos neuroinflamatorios y neurodegenerativos. Eso debilita la antigua afirmación de que CB2 es irrelevante para la enfermedad cerebral. Pero no rescata el viejo binario invirtiéndolo. La esquizofrenia no es solo inflamación, y CB2 no es solo un botón antiinflamatorio. Cualquier modelo traslacional serio tiene que tener en cuenta conjuntamente las interacciones neurona-glía, la señalización inmunitaria y la disfunción de circuitos.
Qué no inferir
- No es prueba de beneficio Que CB1 sea una diana farmacológica no significa que el consumo de cannabis proteja contra la esquizofrenia o la psicosis.
- No refuta el riesgo Los estudios mecanísticos de receptores no anulan la preocupación epidemiológica por la exposición a alto THC en personas vulnerables.
- No son intercambiables Los endocannabinoides, fitocannabinoides y ligandos sintéticos no deben tratarse como el mismo tipo de exposición.
- No es una aprobación general Las aprobaciones existentes de la FDA no resuelven las cuestiones de seguridad psiquiátrica o eficacia para el uso amplio de cannabinoides.
Lo que los lectores no deben inferir de los estudios mecanísticos
Primero, los lectores no deben inferir que, porque CB1 es una diana terapéutica, el consumo de cannabis es por tanto protector frente a la esquizofrenia o la psicosis. La lógica fármaco-diana no funciona así. Los receptores beta son dianas farmacológicas; eso no significa que cualquier estimulante sea un tratamiento. Los receptores opioides son dianas farmacológicas; eso no significa que toda agonía opioide sea beneficiosa. El mismo principio se aplica aquí.
Segundo, los estudios mecanísticos no eliminan la preocupación epidemiológica. La investigación sobre sesgo del receptor y diseño de ligandos convive, no se opone, a la evidencia de que la exposición a cannabis puede empeorar el riesgo de psicosis en algunas personas, especialmente con productos con alto contenido de THC, inicio temprano o consumo intenso. Un mecanismo de receptor puede explicar por qué algunos compuestos podrían diseñarse para obtener mejores resultados. No justifica afirmaciones públicas amplias de que “la activación de CB1 es buena para la esquizofrenia”.
Tercero, los lectores no deben tratar a los endocannabinoids, los fitocannabinoids y los ligandos sintéticos como intercambiables. La anandamida no es THC. Un modulador de CB1 restringido periféricamente no es cannabis fumado. Un compuesto en investigación de baja eficacia y sesgo de vía no es un producto de consumo de alta exposición. Estas categorías importan porque el comportamiento del receptor depende de la clase de ligando, la dosis, la vía, el momento y la exposición tisular.
Por último, la existencia de medicamentos relacionados con cannabis aprobados no resuelve las cuestiones psiquiátricas. A fecha de 2025, la FDA señala que se ha aprobado un producto farmacéutico derivado de cannabis y tres productos farmacéuticos relacionados con cannabis. Ese hecho demuestra que la farmacología cannabinoid puede convertirse en medicina bajo condiciones definidas. No significa que una estimulación no refinada del receptor sea segura, eficaz o apropiada para la esquizofrenia.
La conclusión correcta es más estricta y menos reconfortante. La biología de CB1 es relevante para la psicosis porque el receptor está profundamente integrado en circuitos cerebrales que modelan la saliencia, la cognición y la inhibición. Esa relevancia apoya un diseño cuidadoso de fármacos, no una extrapolación casual. En la investigación sobre esquizofrenia, la farmacología de receptores es al mismo tiempo un mapa de posibilidades y de riesgos.
Lo que la ciencia de receptores significa para los futuros medicamentos cannabinoid
La medicina cannabinoid se está alejando de la vieja idea de que un fármaco útil solo necesita “actuar sobre CB1” o “actuar sobre CB2”. Esa abreviatura siempre fue incompleta. Ahora es activamente engañosa. CB1 y CB2 no son simples interruptores de encendido y apagado divididos limpiamente entre cerebro e sistema inmunitario. Son receptores con distribuciones superpuestas pero desiguales entre tipos celulares, conformaciones dinámicas, preferencias de acoplamiento distintas y roles cambiantes según el estado de la enfermedad. Eso importa porque el mismo ligando puede parecer terapéutico en un tejido, intoxicante en otro e ineficaz en un ensayo clínico si la población con la enfermedad es demasiado amplia o el esquema de dosificación induce desensibilización.
Las apuestas para la salud pública no son menores. La Organización Mundial de la Salud estimó que 200 millones de personas usaron cannabis en 2019, lo que equivale al 4% de la población mundial de 15 a 64 años. Sin embargo, la FDA de EE. UU. sigue señalando que, a fecha de 2025, solo se han aprobado un producto medicinal derivado de cannabis y tres productos medicinales relacionados con cannabis. Esa brecha entre una exposición generalizada y terapéuticas aprobadas limitadas es una de las razones por las que la farmacología de receptores importa tanto. Otra es la carga de enfermedad: la epilepsia afecta a unos 50 millones de personas en todo el mundo, la esquizofrenia a unos 24 millones y el dolor crónico afecta a casi 1 de cada 5 adultos en Estados Unidos. Si la biología de los cannabinoid va a dar lugar a mejores medicamentos, no ocurrirá mediante una activación burda de receptores.
El trabajo fundacional de Raphael Mechoulam, Lumír Hanuš y Allyn Howlett estableció el campo moderno al identificar la señalización de endocannabinoid y la farmacología de receptores. La siguiente fase es diferente. Ya no se trata tanto de demostrar que existen CB1 y CB2, sino de aprender a dirigirlos con precisión.
| Concepto de diseño | Qué pretende mejorar | Por qué el artículo dice que importa |
|---|---|---|
| Selectividad de subtipo | Preferir CB1 o CB2 con mayor precisión | El subtipo por sí solo no garantiza eficacia ni seguridad |
| Ajuste de la eficacia | Evitar una activación del receptor demasiado intensa | Los distintos estados activos pueden cambiar el beneficio, la tolerancia y los efectos adversos |
| Señalización sesgada | Favorecer una vía intracelular sobre otra | Puede ayudar a separar los efectos útiles de las limitaciones |
| Modulación alostérica | Ajustar la respuesta del receptor a los ligandos endógenos | Podría conservar una señalización espacial y temporal más normal |
| Restricción tisular | Limitar la exposición a órganos seleccionados o evitar la entrada al cerebro | Útil cuando predominan los efectos adversos centrales |
Selectividad, eficacia, modulación alostérica y sesgo
La lección más importante de la ciencia de receptores reciente es que “unión” no es lo mismo que “efecto”. Un ligando puede preferir CB1 frente a CB2, o viceversa, pero eso es solo el comienzo. También puede diferir en eficacia, es decir, en cuán intensamente estabiliza estados activos del receptor. Puede favorecer la señalización Gi/o frente al reclutamiento de beta-arrestina, o promover más la internalización del receptor que la señalización sostenida en la superficie. Esas diferencias pueden cambiar drásticamente los resultados.
La revisión de 2026 de Frontiers in Chemical Biology sobre la estructura de receptores cannabinoid deja este punto claro: la selectividad entre subtipos de CB1 y CB2 depende de diferencias estructurales a nivel del receptor que moldean la unión del ligando, la eficacia de señalización y la regulación del receptor. Por eso cannabinoid químicamente similares no se comportan de manera idéntica. Un fitocannabinoid, un endocannabinoid como la anandamida y un ligando sintético pueden contactar con la misma familia de receptores y, aun así, producir perfiles downstream distintos. El diseño de fármacos que ignora la eficacia y el sesgo de señalización ya va por detrás de la evidencia.
CB1 es el caso más claro. A menudo se discute como el receptor responsable de los efectos psicoactivos, lo cual es cierto pero incompleto. CB1 también regula la transmisión sináptica, el procesamiento del dolor, el apetito, los circuitos del estrés y la señalización cortical. El artículo de American Journal of Psychiatry de 2025 sostiene que la señalización sesgada de CB1 es una estrategia terapéutica plausible para la esquizofrenia. Ese es un cambio llamativo en el campo. La esquizofrenia afecta aproximadamente a 24 millones de personas en todo el mundo, y el artículo no trata a CB1 como un receptor que deba evitarse por completo. En cambio, lo trata como una diana cuyas consecuencias patológicas y terapéuticas pueden depender de qué vías intracelulares favorezca un ligando. Esa es la farmacología clásica de GPCR, ahora aplicada con seriedad a las terapias cannabinoid.
CB2 ha experimentado una reevaluación similar. La revisión de 2026 de Frontiers in Behavioral Neuroscience describe “una actualización en los últimos 3 años” que muestra un interés creciente en la señalización de CB2 en trastornos del sistema nervioso central, especialmente cuando intervienen mecanismos neuroinflamatorios y neurodegenerativos. Eso no significa que CB2 sea una diana antiinflamatoria mágica. Significa que la vieja afirmación de que CB2 es irrelevante para el cerebro ya no encaja con la literatura. La expresión puede ser baja en condiciones basales y aumentar en poblaciones celulares concretas o en contextos de enfermedad. Por tanto, un fármaco dirigido a CB2 puede depender en gran medida del momento, la patología y el estado tisular.
La modulación alostérica podría convertirse en una de las formas más útiles de aprovechar estas distinciones. Los agonistas ortostéricos a menudo impulsan una activación amplia del receptor, lo que eleva el riesgo de efectos adversos, tolerancia y escasa selectividad contextual. Los moduladores alostéricos, en cambio, pueden ajustar cómo responden los receptores a ligandos endógenos en lugar de forzar una activación máxima. En principio, eso podría preservar los aspectos espaciales y temporales de la señalización endocannabinoid normal. Es una estrategia atractiva para CB1, donde el agonismo directo a menudo choca con efectos adversos centrales, y para CB2, donde la regulación al alza asociada a la enfermedad puede hacer que la modulación dependiente del contexto resulte más atractiva que la estimulación constante. Pero “atractiva” no significa “demostrada”. Muchas ideas alostéricas que parecen elegantes in vitro siguen fracasando cuando entran en juego la reserva de receptores, la heterogeneidad tisular y la farmacocinética.
Por qué la biología estructural y de redes orienta ahora el diseño de fármacos
El centro de gravedad del campo ha pasado del simple mapeo de receptores al análisis dinámico de estados del receptor. Aquí es donde importa la biología estructural. La revisión de 2026 de Frontiers y el reciente estudio indexado en PubMed sobre el mecanismo dinámico de la selectividad de subtipo empujan contra el modelo llave-cerradura. La selectividad no es solo cuestión de que un ligando encaje mejor en un bolsillo del receptor. Los receptores respiran. Los ligandos estabilizan conjuntos de conformaciones. Los endocannabinoid pueden lograr preferencia de subtipo mediante interacciones dinámicas que alteran cómo CB1 o CB2 pasan entre estados inactivos, activos y competentes para señalización.
Ese cambio de perspectiva tiene consecuencias prácticas. Un químico medicinal ya no pregunta solo: “¿Se une este compuesto a CB1 o CB2?” Las mejores preguntas son: ¿qué conformaciones estabiliza? ¿Promueve Gi/o más que beta-arrestina? ¿Desencadena una internalización rápida? ¿Se comporta de forma distinta en neuronas, microglía, hepatocitos o nociceptores periféricos? ¿Puede restringirse a la periferia? ¿Coopera con el tono del ligando endógeno que solo aumenta durante la inflamación o la lesión?
La biología de redes amplía esto aún más. Un análisis de red integrador reciente indexado en PubMed identificó CB1 y CB2 como nodos altamente influyentes dentro del sistema endocannabinoid y vinculó la señalización del receptor con vías metabólicas más amplias. Eso importa porque la farmacología endocannabinoid nunca trata solo de receptores. La síntesis, el transporte y la degradación de anandamida y 2-AG moldean la exposición al receptor. También lo hacen el metabolismo lipídico, los mediadores inflamatorios y la interacción cruzada con otros sistemas de GPCR y canales iónicos. Un ligando que parece limpio en un ensayo recombinante puede comportarse de forma muy distinta en tejido vivo, donde FAAH, MAGL, las vías de prostaglandinas y la densidad local de receptores varían.
Esta visión sistémica también ayuda a explicar por qué la traslación clínica ha sido desigual. Algunos fracasos probablemente se debieron a compuestos defectuosos. Otros, a modelos de enfermedad excesivamente simplificados. Si un ensayo recluta pacientes cuyos síntomas comparten una etiqueta pero no una firma endocannabinoid, el resultado medio puede parecer decepcionante aunque un subgrupo definido biológicamente pudiera beneficiarse. El éxito futuro dependerá no solo de la química, sino de la estratificación: qué pacientes, qué fase de la enfermedad, qué tejidos, qué estado del receptor, qué tono endógeno.
Expectativas realistas para la próxima generación de terapias
La próxima generación de medicamentos cannabinoid probablemente será más estrecha, más selectiva y menos romántica de lo que sugiere el debate público. Eso es una buena noticia. Los mejores fármacos suelen surgir cuando un campo abandona las explicaciones de talla única.
Ya se vislumbran algunos avances. La restricción periférica de CB1 sigue siendo atractiva para indicaciones de dolor o metabólicas porque busca reducir los efectos adversos centrales. Los compuestos dirigidos a CB2 siguen siendo plausibles para enfermedades inflamatorias, neuropáticas y neuroinmunes, especialmente si la expresión asociada a la enfermedad crea ventanas terapéuticas. Los ligandos sesgados de CB1 pueden resultar útiles en enfermedad neuropsiquiátrica si logran separar los efectos deseados sobre circuitos de la intoxicación, la ansiedad o la tolerancia no deseadas. Los moduladores alostéricos pueden ofrecer un control más sutil que los agonistas directos. Los enfoques dirigidos a enzimas que alteran indirectamente el tono endocannabinoid aún pueden ser valiosos, aunque con sus propios riesgos y sin eludir la complejidad del receptor.
Aun así, el realismo es esencial. La selectividad no garantiza el éxito. Un agonista de CB2 muy selectivo puede fracasar si la expresión de CB2 en la enfermedad diana es demasiado variable o si la señalización downstream no es el verdadero impulsor de los síntomas. El sesgo medido en un ensayo puede no predecir el sesgo en tejido humano. Las instantáneas estructurales son poderosas, pero son instantáneas. La restricción tisular puede reducir algunas responsabilidades a costa de crear otras. Y los fitocannabinoid no deben tratarse como sustitutos de toda la farmacología cannabinoid; los endocannabinoid, los compuestos de origen vegetal y los ligandos sintéticos pueden producir comportamientos de receptor muy distintos según la dosis, el momento y el contexto de señalización.
Así que la previsión sobria es esta: la medicina cannabinoid futura probablemente mejorará al ser más precisa, no al activar de forma más amplia. Las vías más creíbles combinan selectividad de receptor, sesgo de señalización, control alostérico, direccionamiento tisular y mejor estratificación de pacientes. Lo que se sabe es sustancial: CB1 y CB2 son nodos de señalización dinámicos, el estado estructural importa y el contexto de la enfermedad importa. Lo que sigue siendo incierto es si esa claridad mecanística se traducirá en beneficios clínicos reproducibles en dolor, enfermedad psiquiátrica, epilepsia, neurodegeneración y enfermedad inflamatoria. El campo sigue siendo médicamente importante porque la necesidad no cubierta es grande, la biología es real y el modelo simplista anterior por fin ha dado paso a una farmacología capaz de explicar tanto la promesa como la decepción.
Referencias
- [1]Cannabis (marijuana). WHO Fact Sheet, 2024. https://www.who.int/news-room/fact-sheets/detail/cannabis-(marijuana)
- [2]Epilepsy. WHO Fact Sheet, 2024. https://www.who.int/news-room/fact-sheets/detail/epilepsy
- [3]Schizophrenia. WHO Fact Sheet, 2022. https://www.who.int/news-room/fact-sheets/detail/schizophrenia
- [4]FDA Regulation of Cannabis and Cannabis-Derived Products, Including Cannabidiol (CBD). FDA Public Health Focus, 2025. https://www.fda.gov/news-events/public-health-focus/fda-regulation-cannabis-and-cannabis-derived-products-including-cannabidiol-cbd
- [5]The Health Effects of Cannabis and Cannabinoids: The Current State of Evidence and Recommendations for Research. National Academies Press, 2017. https://nap.nationalacademies.org/catalog/24625/the-health-effects-of-cannabis-and-cannabinoids-the-current-state