






Dlaczego CB1 i CB2 mają znaczenie w nauce o cannabis
GPCR Receptor sprzężony z białkiem G: receptor błonowy, który zmienia kształt po związaniu ligandu i przekazuje sygnał przez wewnątrzkomórkowych partnerów, takich jak białka G i beta-arrestyny.
Co determinuje odpowiedź receptora
- Lokalizacja Tkanka, typ komórki i położenie subkomórkowe kształtują odpowiedź.
- Tożsamość ligandu Endokannabinoidy, fitokannabinoidy i ligandy syntetyczne nie wywołują identycznych stanów sygnalizacyjnych.
- Dostępne partnerzy Różne komórki oferują różne białka G, kinazy i beta-arrestyny.
- Wzorzec ekspozycji Czas trwania sygnału i powtarzana stymulacja wpływają na desensytyzację i internalizację.
- Bias szlakowy Ligand może faworyzować sygnalizację przez białka G, rekrutację beta-arrestyn lub inne odpowiedzi.
W powszechnym skrócie myślowym biologia cannabinoidów od dawna sprowadzana jest do zgrabnego podziału: CB1 wyjaśnia „high”, a CB2 odpowiada za zapalenie gdzieś poza mózgiem. Takie ujęcie jest zbyt uproszczone, by było użyteczne. CB1 i CB2 są receptorami sprzężonymi z białkiem G, czyli GPCR, i podobnie jak inne GPCR nie działają jak proste przełączniki „włącz/wyłącz”. Przekazują sygnały pochodzące z endocannabinoidów wytwarzanych przez organizm, fitocannabinoidów z Cannabis sativa oraz syntetycznych ligandów projektowanych w laboratoriach, przekształcając je w zmieniające się odpowiedzi komórkowe. To, jaka odpowiedź się pojawi, zależy od miejsca położenia receptora, od tego, jaki ligand się z nim wiąże, jakie białka sygnałowe są dostępne, jak długo receptor jest stymulowany oraz od tego, czy zostaje on skierowany ku sygnalizacji przez białka G, rekrutacji β-arrestyny, desensytyzacji czy internalizacji.
To ma znaczenie, ponieważ nauka o cannabis nie dotyczy wyłącznie odurzenia. Dotyczy także przewlekłego bólu, padaczki, sygnalizacji immunologicznej, neurodegeneracji, ryzyka psychiatrycznego oraz tego, dlaczego tak wiele programów lekowych opartych na cannabinoidach wyglądało obiecująco przedklinicznie, a następnie potykało się w badaniach z udziałem ludzi. Stawka jest wysoka. World Health Organization oszacowała, że w 2019 roku cannabis używało 200 milionów osób, czyli około 4% światowej populacji w wieku 15–64 lata. Padaczka dotyka około 50 milionów ludzi na świecie. Schizofrenia dotyczy około 24 milionów. A jednak według stanu na 2025 rok U.S. FDA odnotowuje dopuszczenie jednego produktu leczniczego pochodzącego z cannabis i trzech produktów leczniczych związanych z cannabis. Ta przepaść między ogromną ekspozycją a ograniczoną liczbą zatwierdzonych terapii jest jednym z powodów, dla których biologia receptorów ma tak duże znaczenie.
| Cecha | CB1 | CB2 |
|---|---|---|
| Typowy skrót | „receptor mózgowy” | „receptor immunologiczny” |
| Korekta w artykule | Przeważa w ośrodkowym układzie nerwowym, ale występuje też obwodowo | Wzbogacony w układ immunologiczny, ale niepozbawiony znaczenia dla mózgu |
| Przykładowe wymienione funkcje | Percepcja, pamięć, kontrola motoryczna, nocycepcja | Sygnalizacja cytokinowa, migracja komórek, role neurozapalne |
| Interpretacja | Zależna od obwodu i stanu | Zależna od typu komórki i stanu chorobowego |
Dlaczego biologia receptorów wyjaśnia więcej niż etykiety roślinne
Etykiety takie jak „indica”, „sativa”, a nawet „THC-dominant” i „CBD-dominant” mówią tylko część prawdy, ponieważ to receptory, a nie marketingowe kategorie roślin, są najbliżej mechanizmu działania. Δ9-tetrahydrocannabinol (THC) jest częściowym agonistą CB1 i CB2, ale końcowy efekt THC nie jest stały. W neuronach korowych bogatych w CB1 może on hamować uwalnianie neuroprzekaźników i zmieniać percepcję, pamięć oraz kontrolę motoryczną. W obwodowych szlakach czuciowych ta sama rodzina receptorów może kształtować nocycepcję. W komórkach układu immunologicznego aktywacja CB2 może zmieniać sygnalizację cytokinową lub migrację komórek. Ta sama rodzina. Inne skutki.
Prosta zasada, że CB1 jest tylko mózgowy, a CB2 tylko immunologiczny, jest zbyt uproszczona dla współczesnej biologii receptorów.Strong evidence
Stara reguła — CB1 w mózgu, CB2 w komórkach układu immunologicznego — opierała się na rzeczywistym wzorcu, ale źle zniosła próbę czasu. Rozmieszczenie ma charakter gradientowy i zależy od typu komórki, a nie binarne. CB1 jest silnie eksprymowany w wielu regionach centralnego układu nerwowego, zwłaszcza na zakończeniach presynaptycznych, ale występuje również w tkankach obwodowych. CB2 jest mocno związany z funkcją immunologiczną, ale twierdzenie, że nie ma znaczenia dla mózgu, nie da się już obronić. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience argumentował, że sygnalizacja CB2 zyskała uwagę w zaburzeniach centralnego układu nerwowego, szczególnie poprzez mechanizmy neurozapalne i neurodegeneracyjne, i opisał to jako „aktualizację z ostatnich 3 lat”. Ta aktualizacja ma znaczenie. Jeśli CB2 uczestniczy w patologii ośrodkowej w pewnych warunkach, to leków ukierunkowanych na CB2 nie można rozumieć wyłącznie jako narzędzi obwodowych.
Struktura pogłębia ten obraz. Przegląd z 2026 roku w Frontiers in Chemical Biology wyjaśnił, że selektywność ligandów względem „CB1 i CB2” wynika ze структурnych różnic na poziomie receptorów, które wpływają na sposób wiązania, skuteczność działania i regulację receptora. Mówiąc prościej, drobne zmiany chemiczne mogą ukierunkować ligand na jeden podtyp receptora albo na jedną ścieżkę sygnałową, co pomaga wyjaśnić, dlaczego dwa cannabinoidy, które na papierze wyglądają podobnie, mogą wywoływać lub wykazywać bardzo różne efekty in vivo. Badanie indeksowane w PubMed z 2025/2026 roku dotyczące selektywności podtypowej poszło dalej, pokazując, że selektywność endocannabinoidów jest związana z dynamicznymi zmianami konformacji receptora, a nie z sztywnym modelem „zamek-klucz”. Receptor się porusza. Ligand stabilizuje jedne stany bardziej niż inne. Biologia podąża za tymi stanami.
Od fitocannabinoidów do sygnalizacji endocannabinoidowej
{kind=link}
Sygnalizacja wsteczna Wzorzec sygnalizacji synaptycznej, w którym komórka postsynaptyczna uwalnia przekaźnik przemieszczający się wstecznie, aby działać na receptory presynaptyczne.
Sekwencja odkryć w skrócie
- 1 THC wyizolowano i określono jego strukturę.
- 2 W tkance mózgowej wykazano istnienie swoistych miejsc wiązania kannabinoidów.
- 3 CB1 sklonowano jako GPCR.
- 4 CB2 zidentyfikowano w tkankach związanych z układem immunologicznym.
- 5 Anandamid, a następnie 2-AG, ustanowiły endogenny system sygnalizacyjny.
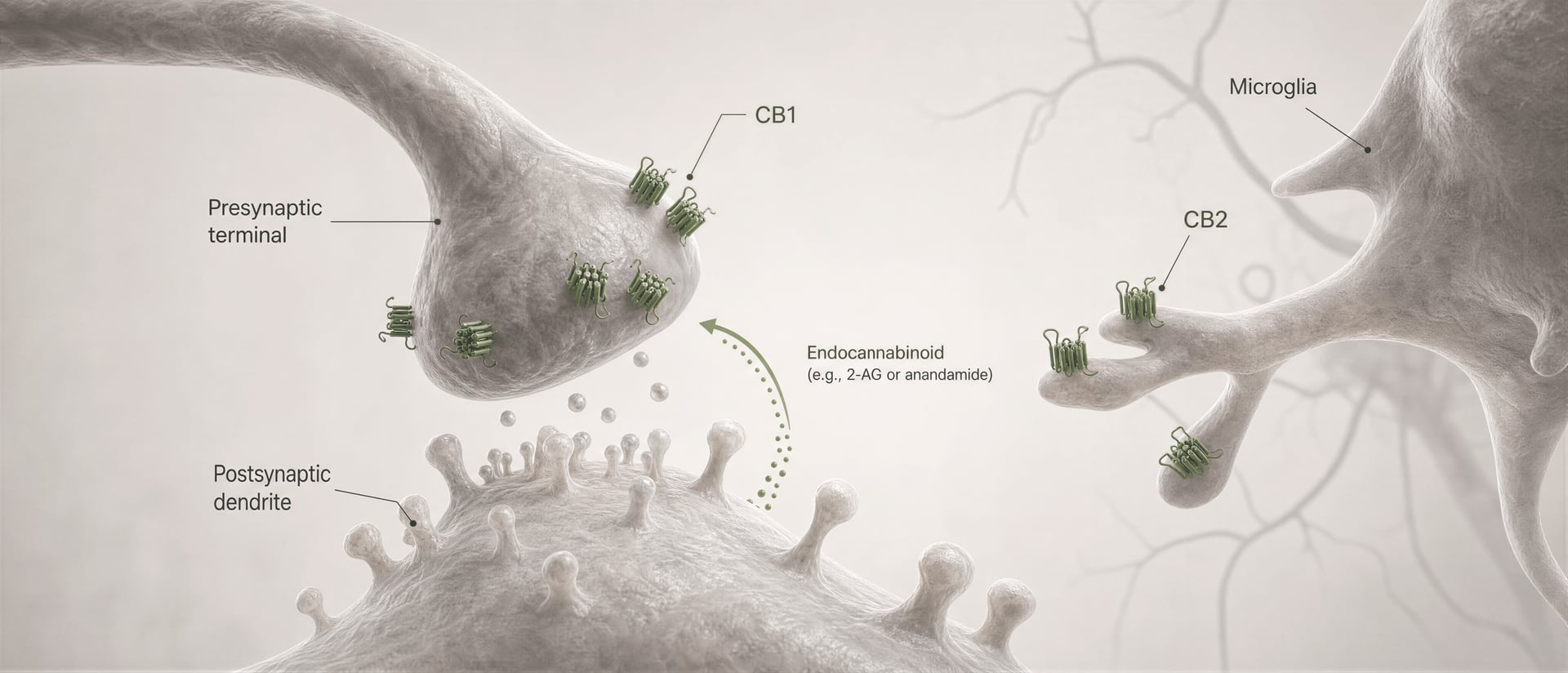
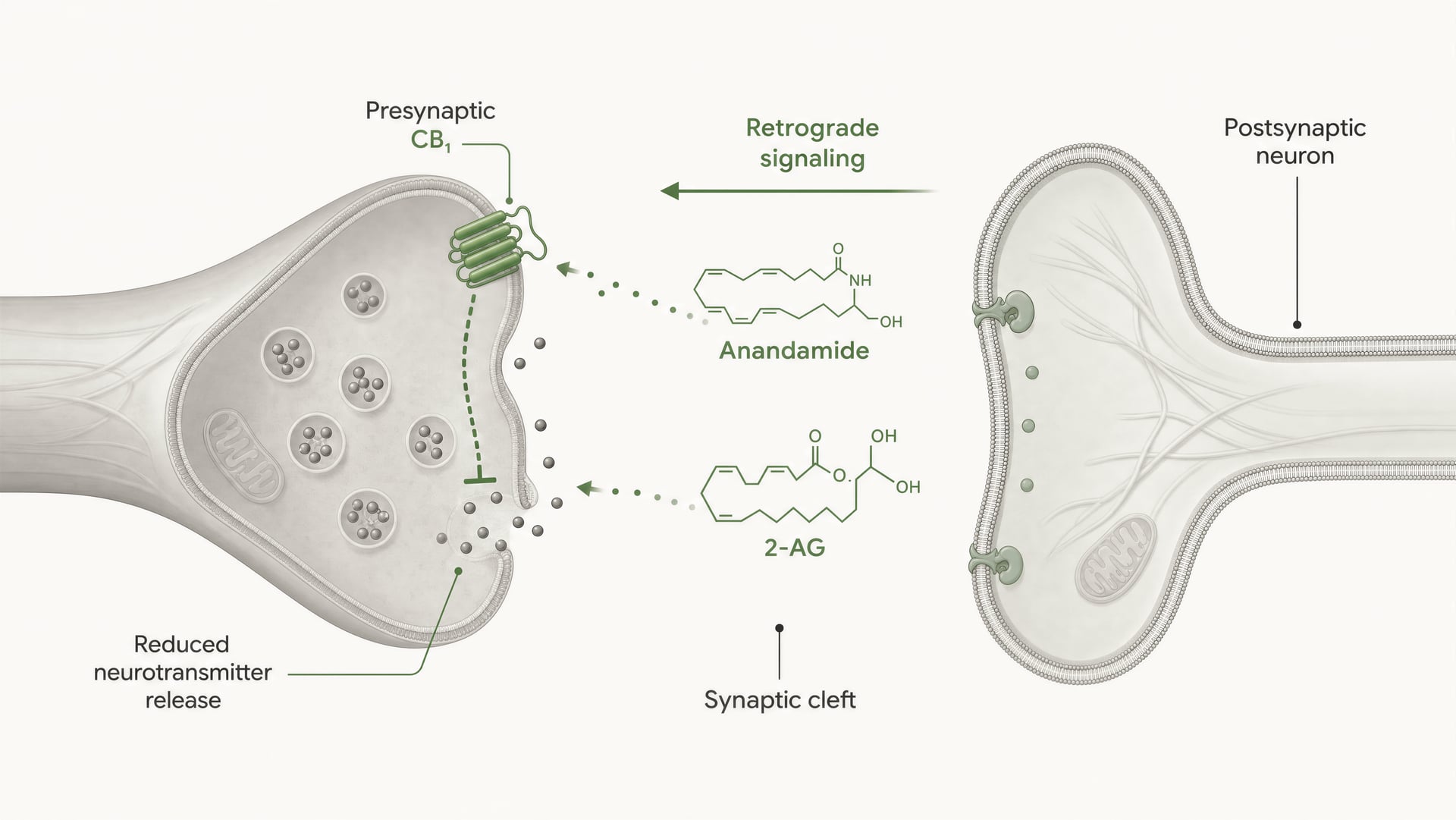
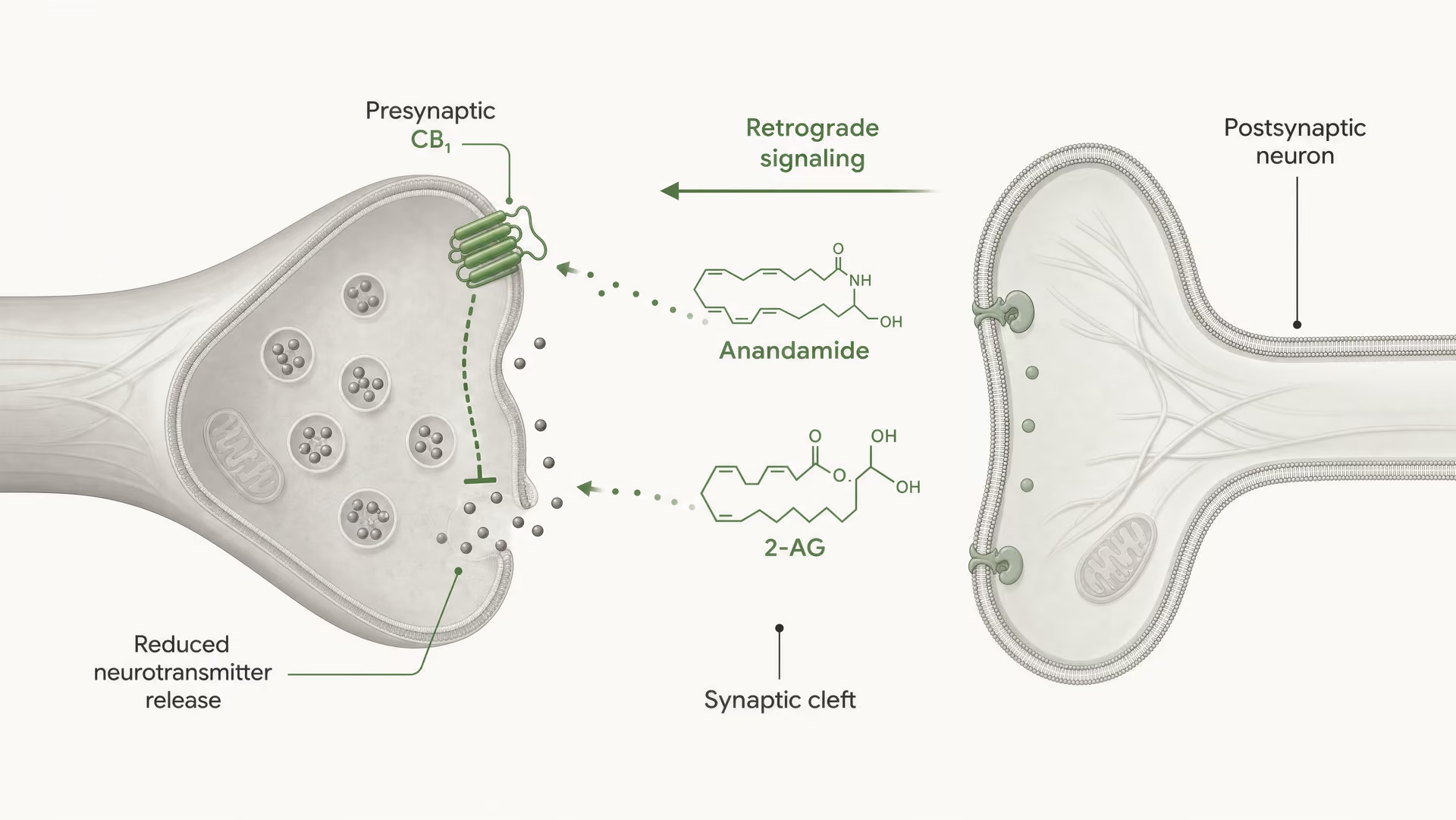
Receptory cannabinoidowe nie zostały odkryte dlatego, że organizm ewoluował z myślą o cannabis. Sekwencja wydarzeń była odwrotna. Prace Allyn Howlett i współpracowników miały kluczowe znaczenie dla zdefiniowania farmakologii receptorów cannabinoidowych, a późniejsze odkrycie anandamidu przez Raphaela Mechoulama i Lumíra Hanuša pomogło ustalić, że ludzie wytwarzają własne cząsteczki sygnałowe podobne do cannabinoidów. Anandamid i 2-arachidonyloglicerol, zwykle skracany do 2-AG, są głównymi endocannabinoidami. Są wytwarzane „na żądanie”, a nie magazynowane w pęcherzykach jak klasyczne neuroprzekaźniki, i często przekazują sygnał w kierunku wstecznym: komórka postsynaptyczna wytwarza endocannabinoid, który przemieszcza się wstecz przez synapsę, aby aktywować presynaptyczny CB1 i zmniejszyć dalsze uwalnianie neuroprzekaźnika.
To zasadniczo inny obraz niż „substancja z cannabis trafia do receptora”. Sygnalizacja endocannabinoidowa jest lokalna, przejściowa i ściśle regulowana przez enzymy syntezy i degradacji. Fitocannabinoidy wchodzą do tego układu z zewnątrz. Ligandy syntetyczne mogą oddziaływać na niego jeszcze silniej lub bardziej selektywnie. W rezultacie ten sam receptor może zostać pobudzony przez krótkotrwały endogenny impuls, powoli wchłaniany doustny fitocannabinoid albo wysokoefiktywny syntetyczny agonista o zupełnie innych zagrożeniach bezpieczeństwa.
Ta różnica jest jednym z powodów, dla których z samej nazwy receptora nie można wywnioskować działania odurzającego. Zależy ono od skuteczności ligandu, dawki, drogi podania, czasu oraz kontekstu tkankowego. THC działające na CB1 jest centralne dla efektów psychoaktywnych, tak, ale fakt ten nie redukuje CB1 do „receptora psychoaktywności”. Nie czyni też z CB2 prostego regulatora przeciwzapalnego. Artykuł z 2025 roku w American Journal of Psychiatry dotyczący biasowanego sygnalizowania przez CB1 dokładnie ten szerszy punkt podnosił, argumentując, że ligandy ukierunkowane na CB1 z preferencją dla określonych szlaków sygnałowych mogą stanowić strategię terapeutyczną w schizofrenii. Ta propozycja łączy naukę o cannabinoidach z szerszą ideą GPCR: jeśli jeden ligand faworyzuje korzystne odgałęzienia sygnalizacji, unikając jednocześnie tych związanych z działaniami niepożądanymi, działanie leku może zostać oddzielone od brutalnej aktywacji receptora. To, czy ta obietnica się utrzyma klinicznie, pozostaje nierozstrzygnięte, ale argument mechanistyczny jest mocny.
Co w tym artykule oznaczają rozmieszczenie, sygnalizacja i cele lekowe
W tym artykule rozmieszczenie oznacza coś więcej niż mapę narządów. Obejmuje gęstość receptorów, typ komórki, lokalizację subkomórkową, stan chorobowy i zmiany w czasie. Receptor eksprymowany na zakończeniach GABA-ergicznych może wywoływać inne skutki obwodowe niż ten sam receptor na zakończeniach glutaminergicznych. Receptor zwiększony w przebiegu zapalenia nie jest równoważny swojemu stanowi wyjściowemu. Rozmieszczenie jest dynamiczne.
Sygnalizacja oznacza wewnątrzkomórkowe konsekwencje pobudzenia receptora. W przypadku CB1 i CB2 obejmuje to sprzężenie z białkami G z rodziny Gi/o, hamowanie cyklazy adenylanowej, modulację kanałów jonowych, zmiany kaskad kinaz, rekrutację β-arrestyny, desensytyzację receptora oraz internalizację. Obejmuje również modulację allosteryczną i agonizm obciążony, w którym ligandy mogą faworyzować jedne wyjścia sygnałowe kosztem innych. To nie są akademickie przypisy. Często to właśnie różnica między analgezją, sedacją, tolerancją, dysforią a nieudanym badaniem klinicznym.
Cele lekowe oznaczają receptory rozważane jako punkt uchwytu interwencji, a nie gwarantowane historie sukcesu. Selektywne ukierunkowanie na CB1 może zmniejszać niektóre działania poza celem, ale nadal napotykać centralne działania niepożądane. Selektywne ukierunkowanie na CB2 może omijać część zagrożeń związanych z odurzeniem, ale selektywność nie gwarantuje skuteczności w złożonych chorobach człowieka. Prace z zakresu biologii systemowej jasno to pokazują. Analiza sieci integracyjnych z 2025/2026 roku indeksowana w PubMed wskazała CB1 i CB2 jako wysoce wpływowe węzły w układzie endocannabinoidowym i powiązała ich sygnalizację z szerszymi szlakami metabolicznymi. Innymi słowy, receptory te znajdują się wewnątrz większych sieci. Naciśnij jeden węzeł, a poruszą się inne szlaki.
Taka jest teza tego artykułu. CB1 i CB2 są zależnymi od kontekstu węzłami sygnałowymi. Nie statycznymi przełącznikami. Nie zwykłymi etykietami dla „mózgu” i „układu odpornościowego”. Jeśli nauka o cannabis ma wyjaśnić, dlaczego jeden związek wygląda na terapeutyczny w jednym kontekście, odurzający w innym, a rozczarowujący w klinice, musi zacząć na poziomie receptora i pozostać tam wystarczająco długo, by śledzić biologię tam, dokąd ona rzeczywiście prowadzi.
Krótka historia odkrycia receptorów cannabinoidowych
Zanim zidentyfikowano receptory cannabinoidowe, nauka o cannabis była przede wszystkim historią chemii. Badacze mogli izolować związki roślinne, porównywać surowe efekty behawioralne u zwierząt i spierać się o moc działania, ale nie potrafili jeszcze wyjaśnić, w jaki sposób cząsteczka taka jak delta-9-tetrahydrokannabinol, czyli THC, wywołuje swoje efekty z precyzją choćby zbliżoną do poziomu receptorowego. Zmieniło się to pod koniec lat 80. i na początku lat 90. XX wieku. Była to zmiana przełomowa: badania nad cannabis przeszły od katalogowania fitocannabinoidów do analizy interakcji ligand-receptor, sygnalizacji wewnątrzkomórkowej, rozmieszczenia w tkankach, a ostatecznie także do badania endogennego układu lipidowego, dziś nazywanego układem endocannabinoidowym, czyli ECS.
| Rok | Kamień milowy | Osoby wymienione w artykule |
|---|---|---|
| 1964 | Izolacja i określenie struktury THC | Raphael Mechoulam; Yechiel Gaoni |
| 1988 | Swoiste miejsca wiązania kannabinoidów o wysokim powinowactwie w błonach mózgu szczura | Allyn Howlett; William Devane |
| 1990 | Klonowanie CB1 | Lisa Matsuda i współpracownicy |
| 1992 | Identyfikacja anandamidu | William Devane; Lumír Hanuš; Raphael Mechoulam; współpracownicy |
| 1993 | Identyfikacja CB2 | Sean Munro; Kerrie Thomas; M. Abu-Shaar |
| 1995 | Identyfikacja 2-AG przez odrębne grupy | zespół Mechoulama; grupa Sugiury |
Od farmakologii THC do identyfikacji receptorów
Kluczowy wczesny kamień milowy miał miejsce w 1964 roku, kiedy Raphael Mechoulam i Yechiel Gaoni opisali izolację oraz strukturę THC. Osiągnięcie to miało znaczenie, ponieważ dało farmakologom zdefiniowaną cząsteczkę do testowania, a nie zmienny ekstrakt botaniczny. Przez kolejne dwie dekady dziedzina budowała mapę zależności struktura-aktywność opartą na THC i pokrewnych cannabinoidach, lecz nadal trwał spór o mechanizm działania. Część badaczy skłaniała się ku nieswoistym efektom błonowym, ponieważ cannabinoidy są lipofilne. Tę hipotezę coraz trudniej było utrzymać w miarę gromadzenia danych o wiązaniu stereoselektywnym i wysycającym.
Era receptorowa rozpoczęła się właściwie wraz z badaniami wiązania w latach 80. W 1988 roku Allyn Howlett i William Devane opublikowali przełomową pracę w Molecular Pharmacology, wykazując specyficzne miejsca wiązania cannabinoidów o wysokim powinowactwie w błonach mózgu szczura przy użyciu syntetycznego agonisty CP55,940. Wynik ten nie był mglistą sugestią istnienia celu działania. Pokazywał wysycalność, zróżnicowanie regionalne i swoistość farmakologiczną zgodną z obecnością rzeczywistego receptora. Tkanka mózgowa nie reagowała na cannabinoidy tak, jakby po prostu rozpuszczały się one w dwuwarstwie lipidowej i zakłócały wszystko jednocześnie. Istniała selektywność.
Trzy lata później, w 1990 roku, Lisa Matsuda i współpracownicy sklonowali pierwszy receptor cannabinoidowy, dziś nazywany CB1, i opublikowali go w Nature. CB1 został zidentyfikowany jako receptor sprzężony z białkiem G, czyli GPCR, co natychmiast umieściło farmakologię cannabinoidów w obrębie jednej z najważniejszych nadrodzin sygnałowych w biologii. Miało to ogromne znaczenie, ponieważ GPCR nie są jedynie przełącznikami. Przyjmują wiele stanów konformacyjnych, łączą się z różnymi partnerami wewnątrzkomórkowymi, ulegają desensytyzacji, internalizacji i wykazują zależną od ligandu stronniczość sygnałową. Te idee miały stać się centralne znacznie później, ale klonowanie CB1 uczyniło je możliwymi.
CB2 pojawił się szybko potem. W 1993 roku Sean Munro, Kerrie Thomas i M. Abu-Shaar zidentyfikowali drugi receptor cannabinoidowy, CB2, również w Nature, początkowo scharakteryzowany w tkankach związanych z układem odpornościowym. To odkrycie stworzyło trwały skrót myślowy, który przez lata kształtował dziedzinę: CB1 jako „receptor mózgowy” związany z odurzeniem, CB2 jako „obwodowy” lub immunologiczny receptor związany ze stanem zapalnym. Skrót ten był użyteczny, ale nawet wtedy był zbyt uproszczony, a z czasem wyraźnie się zdezaktualizował. Rozmieszczenie obu receptorów zależy od gatunku, typu komórki, stanu aktywacji, kontekstu chorobowego oraz metody badawczej.
Jak CB1 i CB2 zmieniły dziedzinę endocannabinoidów
Gdy zidentyfikowano CB1 i CB2, oczywistym następnym pytaniem było, dlaczego organizm w ogóle posiada receptory dla cannabinoidów pochodzenia roślinnego. Odpowiedź nadeszła w 1992 roku, kiedy William Devane, Lumír Hanuš, Raphael Mechoulam i współpracownicy zidentyfikowali anandamid, formalnie etanoloamid arachidonowy, jako endogenny ligand. Praca opublikowana w Science oznaczała przełom koncepcyjny. Farmakologia cannabis nie dotyczyła już wyłącznie związków egzogennych pochodzących z Cannabis sativa. Dotyczyła natywnego układu sygnalizacji lipidowej.
Drugi główny endogenny ligand, 2-arachidonyloglicerol, czyli 2-AG, został zidentyfikowany w 1995 roku przez odrębne zespoły, w tym grupę Mechoulama oraz grupę Sugiury. Wraz z receptorami i endogennymi ligandami ECS szybko się rozwinął. Badacze zidentyfikowali enzymy syntetyzujące i degradujące, takie jak hydrolaza amidów kwasów tłuszczowych, FAAH, odpowiedzialna za anandamid, oraz lipaza monoacyloglicerolowa, MAGL, odpowiedzialna za 2-AG. Musieli też zmierzyć się z nadal nierozstrzygniętym problemem: w jaki sposób te silnie lipofilne cząsteczki przemieszczają się przez błony i przestrzeń pozakomórkową. W dziedzinie często mówi się o „transporcie”, lecz pojedynczy, wyspecjalizowany transporter endocannabinoidów pozostaje nieuchwytny.
W tym momencie nauka o cannabinoidach przestała być schematem dwóch receptorów i stała się siecią sygnałową. CB1 i CB2 powiązano z białkami Gi/o, hamowaniem cyklazy adenylanowej, modulacją kanałów wapniowych i potasowych oraz hamowaniem uwalniania neuroprzekaźników. Jednak historia nie pozostała tak prosta. Receptory mogły rekrutować beta-arrestyny, ulegać desensytyzacji i internalizacji oraz reagować odmiennie na fitocannabinoidy, endocannabinoidy i ligandy syntetyczne, nawet jeśli formalnie trafiały one w ten sam receptor. Współczesny język GPCR dotyczący agonizmu stronniczego szczególnie dobrze pasuje do cannabinoidów. Artykuł z 2025 roku w American Journal of Psychiatry argumentował, że stronnicza sygnalizacja CB1 jest prawdopodobną strategią terapeutyczną w schizofrenii, zaburzeniu dotykającym około 24 milionów ludzi na świecie według WHO. To bardzo długa droga od dawnego obrazu CB1 jako jedynie receptora wyjaśniającego odurzenie THC.
Historia CB2 również uległa poszerzeniu. Wczesne badania lokowały go głównie w tkankach odpornościowych i było to kierunkowo trafne, lecz późniejsze prace wykazały ekspresję CB2 w mikrogleju oraz w niektórych warunkach także w innych populacjach komórek ośrodkowego układu nerwowego. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience opisywał „aktualizację z ostatnich 3 lat” łączącą sygnalizację CB2 z mechanizmami neurozapalnymi i neurodegeneracyjnymi, jasno pokazując, że CB2 nie można uznać za nieistotny dla mózgu. Najnowsze badania strukturalne poszły jeszcze dalej. Przegląd z 2026 roku w Frontiers in Chemical Biology dotyczący „CB1 i CB2” podkreślał, że selektywność podtypowa zależy od różnic strukturalnych na poziomie receptora, które zmieniają wiązanie, skuteczność i regulację. Niedawne badanie indeksowane w PubMed dotyczące selektywności podtypowej również wskazuje, że selektywność endocannabinoidów jest dynamiczna i kształtowana przez zachowanie konformacyjne, a nie przez prosty model klucza i zamka.
Badacze fundamentów i dlaczego ta historia nadal ma znaczenie
Trzy nazwiska zajmują centralne miejsce w tej historii. Raphael Mechoulam pomógł określić chemiczne i biologiczne podstawy nauki o cannabinoidach, od badań nad strukturą THC po odkrycie endocannabinoidów. Lumír Hanuš był kluczową postacią w identyfikacji anandamidu i późniejszych badań nad endocannabinoidami. Farmakologia receptorowa Allyn Howlett miała decydujące znaczenie dla wykazania, że cannabinoidy działają poprzez swoiste miejsca wiązania w mózgu i konkretne mechanizmy sygnalizacyjne. Bez ich pracy nie byłoby współczesnej dziedziny ECS.
Ta historia nadal ma znaczenie, ponieważ dawne uproszczenia wciąż zniekształcają współczesne debaty. W 2019 roku około 200 milionów ludzi na świecie, czyli około 4% osób w wieku 15–64 lat, używało cannabis według WHO. Jednocześnie FDA podaje, że zatwierdziła jeden produkt leczniczy pochodzący z cannabis oraz trzy produkty lecznicze związane z cannabis. Ekspozycja publiczna jest ogromna. Przekład na praktykę kliniczną jest selektywny i trudny. Historia receptorów wyjaśnia dlaczego. Efekty cannabinoidów zależą od klasy ligandu, stanu receptora, lokalizacji tkankowej, czasu działania i stronniczości szlaku sygnałowego. Zależą też od szerszej sieci. Analiza sieci integracyjnej z lat 2025/2026 zidentyfikowała CB1 i CB2 jako wysoce wpływowe węzły powiązane ze szlakami metabolicznymi, a nie jako izolowane cele.
To jest prawdziwe dziedzictwo odkrycia receptorów. Nie uprościło ono biologii cannabis. Pokazało, dlaczego ta biologia jest bardziej złożona, niż kiedykolwiek zakładał dawny podział na mózg i ciało.
Gdzie występuje CB1: obwody mózgowe, tkanki obwodowe i gradienty funkcjonalne
CB1 zasłużył na swoją reputację głównego psychoaktywnego receptora cannabinoidowego nie bez powodu. Jest obficie obecny w ośrodkowym układzie nerwowym, a prace farmakologiczne Allyn Howlett pomogły ustalić, że THC działa poprzez swoisty, wysycający się układ receptorowy, a nie przez nieswoiste efekty błonowe. Jednak dawny skrót myślowy — CB1 w mózgu, CB2 w komórkach odpornościowych — dziś powoduje więcej zamieszania niż jasności. CB1 jest rzeczywiście silnie wzbogacony w obwodach neuronowych. Jest też obecny w jelitach, wątrobie, tkance tłuszczowej, narządach rozrodczych, tkankach układu sercowo-naczyniowego i szlakach czuciowych, gdzie wpływa na przyjmowanie pokarmu, metabolizm, sygnalizację bólową i funkcje autonomiczne. Rozmieszczenie jest szerokie. Funkcja jest warunkowa.
Ma to znaczenie, ponieważ ekspozycja na cannabinoidy jest powszechna. Światowa Organizacja Zdrowia oszacowała, że w 2019 roku 200 milionów ludzi używało cannabis, czyli około 4% globalnej populacji w wieku 15–64 lat. Ma to również znaczenie, ponieważ farmakologia receptorowa coraz częściej wchodzi do medycyny: FDA podaje, że zatwierdziła jeden produkt leczniczy pochodzący z cannabis oraz trzy produkty lecznicze związane z cannabis. Receptor obecny w tak wielu narządach nie może być sprowadzony do jednej etykiety behawioralnej.
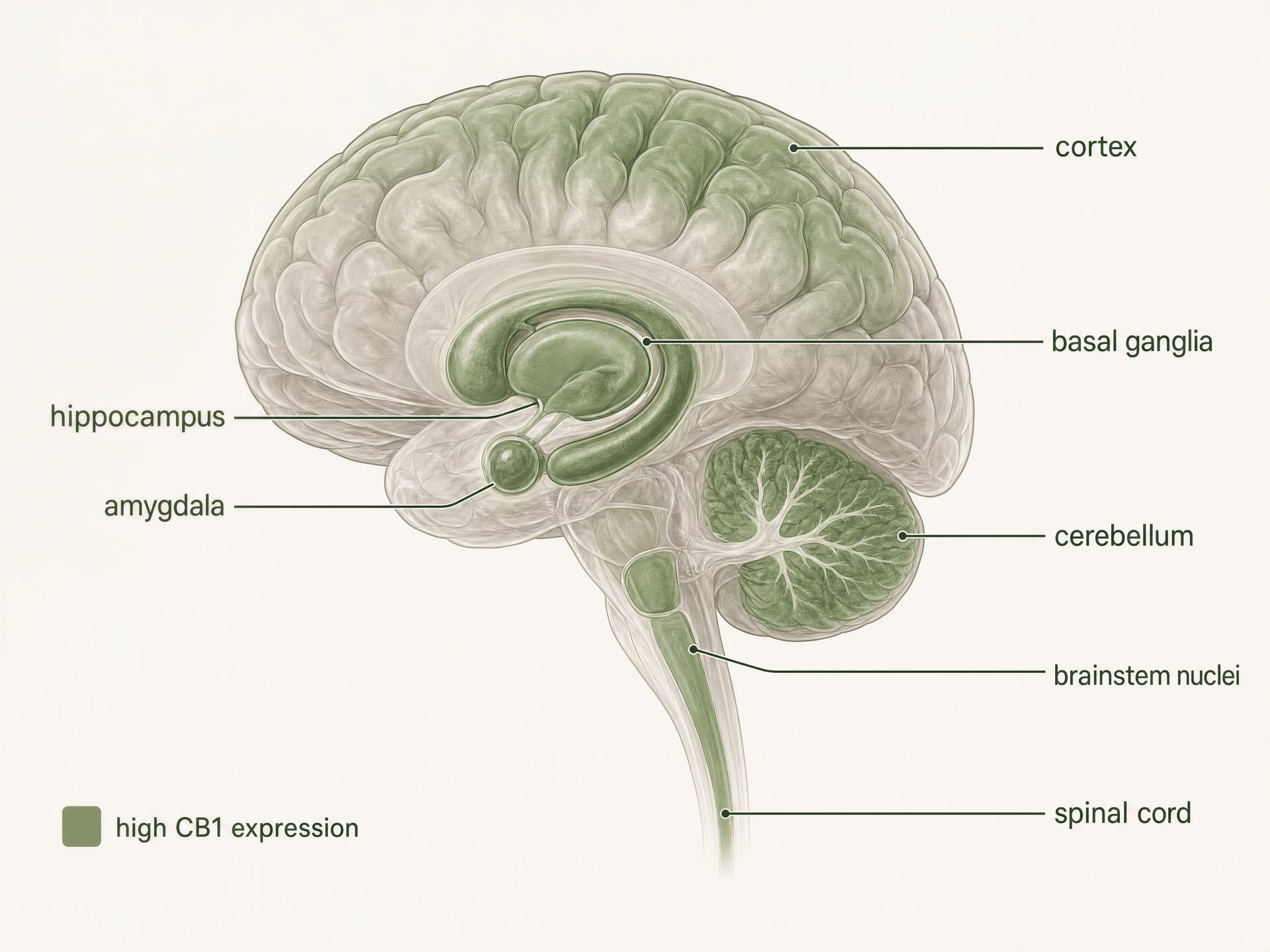
- Ogólny wzorzec
- Jeden z najobficiej występujących GPCR w mózgu ssaków
- Wymienione regiony o dużej gęstości
- Kora mózgowa, hipokamp, ciało migdałowate, jądra podstawy, móżdżek
- Wymienione miejsca związane z bólem
- Istota szara okołowodociągowa, brzuszno-przyśrodkowa część rdzenia przedłużonego, róg tylny rdzenia kręgowego
- Wymienione miejsca obwodowe
- Jelito, wątroba, tkanka tłuszczowa, szlaki rozrodcze, sercowo-naczyniowe, czuciowe
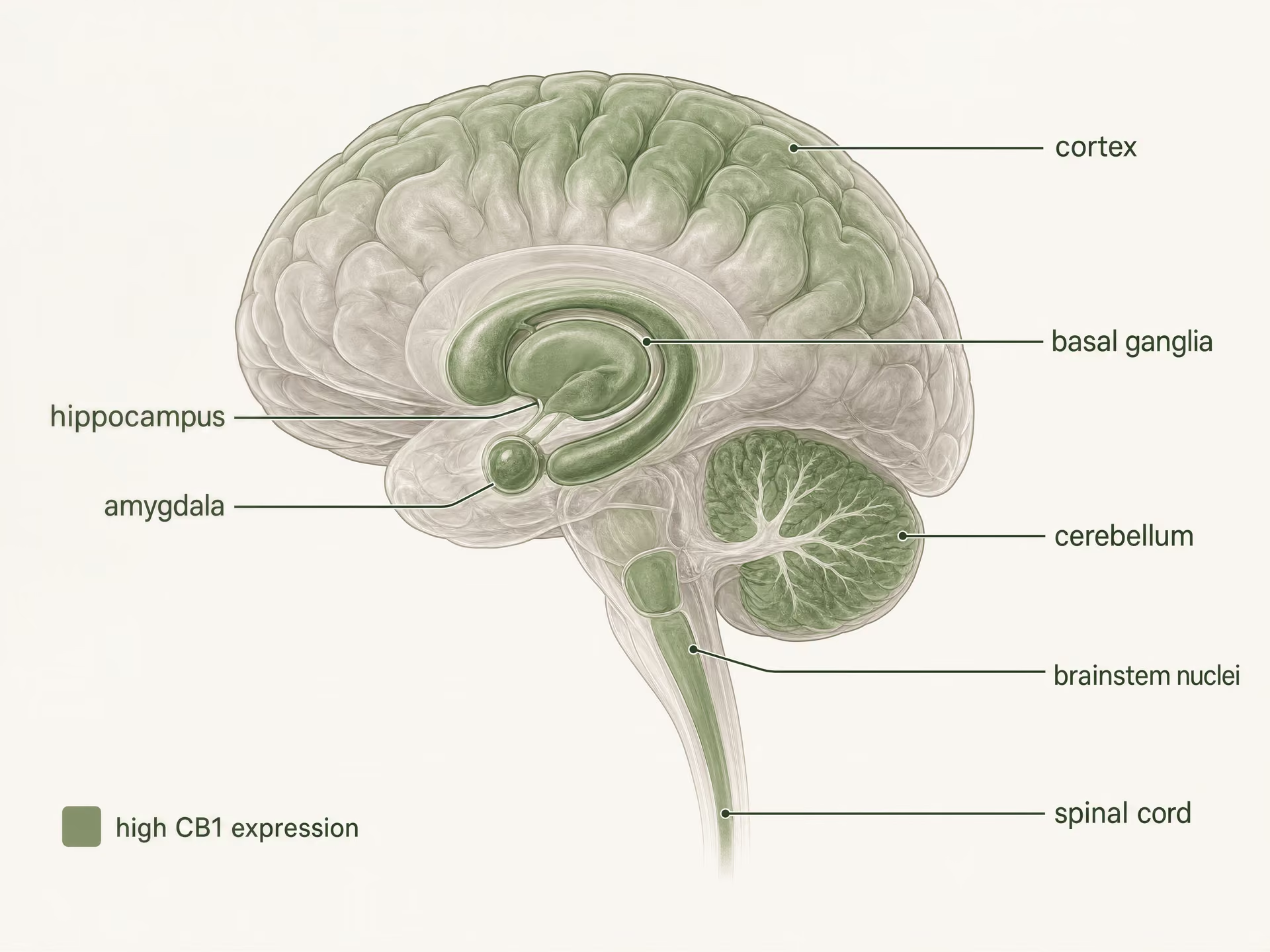
Wysoka ekspresja w ośrodkowym układzie nerwowym
{kind=link}
CB1 jest jednym z najliczniej występujących receptorów sprzężonych z białkiem G w mózgu ssaków. Autoradiografia, hybrydyzacja in situ i mapowanie immunohistochemiczne stworzyły jasny obraz na długo przed współczesnymi badaniami strukturalnymi: wysokie gęstości występują w korze mózgowej, hipokampie, ciele migdałowatym, zwojach podstawy, móżdżku oraz w kilku regionach przetwarzania bólu, z dodatkową ekspresją w jądrach pnia mózgu i na całej długości rdzenia kręgowego. Taki wzorzec zaskakująco dobrze, choć nie idealnie, odpowiada klasycznym efektom THC.
| Położenie CB1 | Bezpośredni efekt synaptyczny | Przykładowy skutek wymieniony w artykule |
|---|---|---|
| Zakończenia interneuronów GABA-ergicznych | Hamuje uwalnianie GABA | Odtłumienie neuronów postsynaptycznych |
| Zakończenia glutaminianergiczne | Hamuje uwalnianie glutaminianu | Osłabienie pobudzenia |
| Obwody jąder podstawy i móżdżku | Zmienia uwalnianie neuroprzekaźników w szlakach ruchowych | Spowolnienie ruchowe, zmienione obwody nawykowe, zaburzona koordynacja |
| Szlaki bólowe | Moduluje przewodnictwo nocyceptywne | Zmiany w zstępującym, wstępującym, zapalnym i afektywnym przetwarzaniu bólu |
W korze mózgowej i hipokampie CB1 występuje w obwodach regulujących uwagę, pamięć roboczą, uczenie wygaszania oraz krótkotrwałą plastyczność synaptyczną. Efekty dotyczące pamięci nie sprowadzają się do prostego schematu „hipokamp równa się zapominanie”. W dużej mierze zależą od tego, które zakończenia aksonalne wykazują ekspresję receptora. CB1 często koncentruje się presynaptycznie, gdzie po aktywacji przez endocannabinoid, takie jak anandamid i 2-arachidonyloglicerol, hamuje uwalnianie neuroprzekaźników; te lipidy sygnałowe, których odkrycie przez Raphaela Mechoulama, Lumíra Hanuša i współpracowników zmieniło całą dziedzinę. Gdy CB1 jest aktywowany na zakończeniach interneuronów GABAergicznych, może znosić hamowanie neuronów niższego rzędu; gdy jest aktywowany na zakończeniach glutaminergicznych, może tłumić pobudzenie. Ten sam receptor, odwrotny wynik sieciowy.
Jądra podstawne i móżdżek wyjaśniają kolejny znany zestaw efektów. Gęsta ekspresja CB1 w prążkowiu, gałce bladej, istocie czarnej pars reticulata oraz w warstwach drobinowych móżdżku wiąże aktywację receptora ze spowolnieniem ruchowym, zmianą obwodów nawyku, zaburzeniem koordynacji oraz, przy niektórych dawkach, efektami przypominającymi katalepsję w modelach zwierzęcych. Jednocześnie fakt, że CB1 występuje skąpo w ośrodkach pnia mózgu kontrolujących układ krążeniowo-oddechowy w porównaniu z receptorami opioidowymi, pomaga wyjaśnić, dlaczego przedawkowanie cannabinoidów zwykle nie wywołuje takiego samego wzorca śmiertelnej depresji oddechowej jak silni agoniści opioidowi. Znaczenie ma lokalizacja. Znaczenie ma też to, czego nie ma.[5]The Health Effects of Cannabis and Cannabinoids: The Current State of Evidence and Recommendations for Research. National Academies of Sciences, Engineering, and Medicine. National Academies Press, 2017. https://nap.nationalacademies.org/catalog/24625/the-health-effects-of-cannabis-and-cannabinoids-the-current-state
Przetwarzanie bólu pokazuje tę samą regionalną logikę. CB1 występuje w istocie szarej okołowodociągowej, brzuszno-przyśrodkowej części rdzenia przedłużonego, rogu tylnym rdzenia kręgowego oraz obwodowych szlakach nocyceptywnych. Daje to receptorowi wiele punktów wejścia do nocycepcji: może on modyfikować wstępujące sygnały bólowe, zstępującą kontrolę bólu, uczulenie zapalne oraz emocjonalne zabarwienie bólu. To jeden z powodów, dla których cannabinoids pozostają obecne w dyskusji o bólu przewlekłym, zwłaszcza że niemal 1 na 5 dorosłych w Stanach Zjednoczonych żyje z bólem przewlekłym, zgodnie z National Academies. Jednak analgezja nie jest gwarantowana tylko dlatego, że CB1 jest obecny. Sedacja, upośledzenie funkcji poznawczych, tolerancja i działania niepożądane ograniczające dawkę często pojawiają się za pośrednictwem sąsiednich obwodów albo tych samych obwodów przy innym poziomie zaangażowania receptora.
Biegunowane sygnalizowanie CB1 może oddzielać pożądane efekty terapeutyczne od niepożądanych efektów psychoaktywnych lub poznawczych.Limited evidence
Białka Gi/o Rodzina białek G, które zwykle zmniejszają aktywność cyklazy adenylanowej i pomagają kontrolować kanały jonowe po aktywacji GPCR.
Nowoczesna biologia receptorów dodaje kolejny poziom. CB1 nie jest prostym przełącznikiem włącz/wyłącz. Głównie sprzęga się z białkami Gi/o, zmniejszając aktywność cyklazy adenylanowej i modulując kanały jonowe, ale może też rekrutować beta-arrestyny, ulegać desensytyzacji i internalizacji oraz wykazywać zależne od ligandu skrzywienie sygnałowe. Artykuł z 2025 roku w American Journal of Psychiatry, argumentujący, że skrzywione sygnałowanie CB1 mogłoby zostać wykorzystane terapeutycznie w schizofrenii, stawia tę kwestię wprost: samo zajęcie receptora jest słabym predyktorem wyniku klinicznego. Ponieważ schizofrenia dotyczy około 24 milionów osób na świecie, atrakcyjna jest możliwość oddzielenia pożądanego sygnalizowania od niepożądanych efektów psychoaktywnych lub poznawczych. Czy takie rozdzielenie da się osiągnąć w praktyce, pozostaje otwartym pytaniem rozwoju leków, a nie ustalonym faktem.
Obwodowy CB1 w jelicie, wątrobie, tkance tłuszczowej i poza nimi
CB1 poza mózgiem nie jest przypisem. To klucz do zrozumienia, dlaczego cannabinoids wpływają na apetyt, nudności, gospodarkę glukozową, metabolizm lipidów i czucie trzewne.
W jelicie CB1 jest eksprymowany w enteric nervous system, przedziałach nabłonkowych oraz szlakach związanych z nerwem błędnym. Aktywacja może spowalniać opróżnianie żołądka, zmieniać motorykę jelit, zmniejszać wymioty i modyfikować sygnalizację między jelitem a mózgiem. Efekty dotyczące apetytu często opisuje się tak, jakby pochodziły wyłącznie z podwzgórzowych ośrodków nagrody i karmienia, lecz obwodowy CB1 współtworzy ten obraz, kształtując bodźce czuciowe i hormonalne jeszcze zanim sygnały dotrą do tych obwodów. Posiłek nie oddziałuje na pusty krajobraz receptorowy; lokalnie zmienia ton endocannabinoidów.
W wątrobie i tkance tłuszczowej CB1 uczestniczy w regulacji metabolicznej, w tym w lipogenezie, wrażliwości na insulinę i magazynowaniu energii. Była to jedna z głównych lekcji ery rimonabantu. Blokowanie CB1 poprawiało masę ciała i parametry metaboliczne, co wspierało pogląd, że nadmierna aktywność sygnalizacji endocannabinoidów przyczynia się do patologii związanej z otyłością. Jednak rimonabant, centralnie aktywny inverse agonist CB1, powodował również poważne działania niepożądane psychiatryczne, w tym depresję i lęk, i został wycofany. Ten epizod często przywołuje się jako porażkę „celowania w CB1”. Trafniej byłoby powiedzieć, że była to porażka określonego rodzaju celowania w CB1: silnej centralnej antagonizacji lub inverse agonism w systemie receptorowym osadzonym w obwodach nastroju i stresu. Wniosek nie brzmi, że obwodowy CB1 jest nieistotny; brzmi on tak, że wzorzec ekspozycji na lek i stan receptora są równie ważne jak sama nazwa receptora.
Adipocyty, hepatocyty, tkanka trzustkowa, mięśnie szkieletowe, tkanki układu sercowo-naczyniowego oraz narządy rozrodcze dopełniają mapę obwodową. Tak samo neurony czuciowe. Integracyjna analiza sieciowa z 2025/2026 roku, indeksowana w PubMed i identyfikująca CB1 oraz CB2 jako bardzo wpływowe węzły w sygnalizacji endocannabinoid, jest tu użyteczna, ponieważ przesuwa uwagę z samej lokalizacji receptora na jego udział w sieciach metabolicznych i sygnałowych. Receptor o umiarkowanej ekspresji w jednej tkance może nadal wywierać duże efekty systemowe, jeśli znajduje się w punkcie wąskim lokalnego sygnalizowania.
Badania strukturalne także utrzymują tę dyskusję w ryzach. Przegląd z 2026 roku w Frontiers in Chemical Biology dotyczący CB1 i CB2 podkreśla, że selektywność ligandu i skuteczność wynikają ze strukturalnych różnic na poziomie receptora, które zmieniają wiązanie, sygnalizację i regulację receptora. Badanie z 2025/2026 roku, indeksowane w PubMed, dotyczące selektywności wobec podtypów, również argumentuje, że dynamika konformacyjna, a nie tylko dopasowanie na zasadzie klucz-zamek, kształtuje to, jak endocannabinoid rozróżniają podtypy receptorów. Ma to znaczenie dla dystrybucji CB1, ponieważ „CB1 w wątrobie” nie oznacza, że THC, anandamid, 2-AG i syntetyczny agonista zrobią tam to samo.
Dlaczego rozmieszczenie nie oznacza jednej, jednolitej funkcji
Największym błędem w mapach receptorowych jest traktowanie ekspresji jako przeznaczenia. Tak nie jest. Wysoka ekspresja mówi, gdzie szukać, nie co się wydarzy.
Po pierwsze, typ komórki zmienia znak efektu. Receptor CB1 na zakończeniu glutaminergicznym może zmniejszać pobudzenie. Ten sam receptor na zakończeniu GABAergicznym może zmniejszać hamowanie. To nie są zamienne wyniki. Po drugie, znaczenie ma lokalizacja synaptyczna. CB1 zwykle występuje presynaptycznie, często aktywowany przez endocannabinoid uwalniane „na żądanie” z neuronów postsynaptycznych, co tworzy retrogradową kontrolę uwalniania neuroprzekaźników. Taki układ sprzyja krótkiej, zależnej od aktywności modulacji, a nie stałej aktywacji receptora.
Po trzecie, znaczenie ma tożsamość ligandu. Endocannabinoid to krótkotrwałe miejscowe przekaźniki. Phytocannabinoid, takie jak THC, przychodzą z zewnątrz układu, często przy wyższej i bardziej długotrwałej ekspozycji niż sygnały endogenne. Ligandy syntetyczne mogą działać jeszcze silniej, z inną skutecznością i innym skrzywieniem sygnałowym. Niektóre silniej promują sygnalizację Gi/o; inne faworyzują rekrutację beta-arrestyn, desensytyzację lub internalizację receptora. Dlatego dwa związki mogą być nazwane agonistami CB1, a mimo to wyraźnie różnić się nasileniem pobudzenia apetytu, zaburzeń pamięci, upośledzenia ruchowego i rozwoju tolerancji.
Po czwarte, lokalna dostępność ligandu zmienia wszystko. Anandamid i 2-AG są syntetyzowane i rozkładane na miejscu, więc ich działanie zależy od aktywności neuronalnej, stanu metabolicznego, ekspresji enzymów i kontekstu zapalnego. Po piąte, sama gęstość receptorów istnieje na gradiencie. Obszar mózgu, etap rozwoju, stan chorobowy i powtarzana ekspozycja na lek wszystkie zmieniają poziom CB1 i odpowiedź na jego aktywację.
Aktualna literatura odchodzi od binarności właśnie z tego powodu. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience odnotowuje aktualizację z ostatnich 3 lat w sposobie rozumienia sygnalizacji receptorów cannabinoid w zaburzeniach OUN, zwłaszcza po uwzględnieniu mechanizmów neurozapalnych i neurodegeneracyjnych. CB1 należy odczytywać z podobną ostrożnością. Jest dominującym receptorem ośrodkowym, ale nie wyłącznie ośrodkowym; receptorem regulującym łaknienie, ale nie tylko tym; celem przeciwbólowym, ale nie czystym przełącznikiem analgezji. Każdy poważny opis dystrybucji CB1 musi myśleć w kategoriach gradientów, obwodów i stanów sygnalizacyjnych, a nie w kategoriach uproszczonej opozycji mózg kontra ciało.
Gdzie występuje CB2: korzenie w układzie odpornościowym i rozszerzająca się mapa OUN
Stare uproszczenie mówiło to jasno: CB1 to receptor mózgowy, CB2 to receptor immunologiczny. Taki schemat pomagał w nauczaniu, ale dziś bardziej zaciemnia niż wyjaśnia. CB2 rzeczywiście wykazuje klasyczne wzbogacenie poza neuronami, zwłaszcza w liniach komórkowych układu odpornościowego i hematopoetycznego, i ten fakt nadal ma znaczenie dla farmakologii. Jednak nowsza literatura, zwłaszcza przegląd z 2026 roku w Frontiers in Behavioral Neuroscience, stawia mocniejsze twierdzenie: o CB2 mówi się obecnie także w kontekście zaburzeń centralnego układu nerwowego, ponieważ jego ekspresja i sygnalizacja stają się bardziej widoczne w mikrogleju, obwodach zapalnych i stanach związanych z uszkodzeniem — to „aktualizacja z ostatnich 3 lat”, która zmieniła sposób mapowania i interpretacji tego receptora. Wniosek nie jest taki, że CB2 nagle stał się wysoko obfitym, neuronalnym receptorem całego mózgu. Tak się nie stało. Wniosek brzmi, że dystrybucję tego receptora trzeba opisywać jako zależną od warunków, specyficzną dla typu komórki i zależną od stanu.
To rozróżnienie ma znaczenie kliniczne. Światowa Organizacja Zdrowia oszacowała, że w 2019 roku cannabis używało 200 milionów osób, czyli 4% globalnej populacji w wieku 15–64 lata. Nawet przy zaledwie niewielkiej liczbie zatwierdzonych leków związanych z cannabinoidami — FDA w 2025 roku wskazywała jeden produkt pochodzenia cannabis i trzy produkty związane z cannabis — lokalizacja receptorów nadal wyznacza, gdzie twórcy leków szukają efektów przeciwzapalnych, przeciwbólowych, neuroprotekcyjnych i psychiatrycznych oraz gdzie spodziewają się działań niepożądanych.
- Klasyczne wzbogacenie
- Komórki układu immunologicznego i krwiotwórczego
- Wymienione typy komórek
- Limfocyty B, limfocyty T, makrofagi, monocyty, komórki NK, neutrofile, komórki tuczne
- Wymienione tkanki kanoniczne
- Śledziona, migdałek, grasica, szpik kostny, krążące komórki układu immunologicznego
- Podkreślone znaczenie dla OUN
- Mikroglej i stany związane z patologią
Klasyczne wzbogacenie w komórkach odpornościowych i hematopoetycznych
CB2 został pierwotnie zidentyfikowany jako podtyp receptora cannabinoidowego o najsilniejszej ekspresji w komórkach związanych z odpornością, a nie z szybką transmisją synaptyczną. To nadal jest właściwy punkt wyjścia. W porównaniu z CB1, który jest silnie reprezentowany w wielu populacjach neuronalnych, CB2 jest klasycznie wzbogacony w komórkach B, komórkach T, makrofagach, monocytach, komórkach NK, neutrofilach, komórkach tucznych oraz innych kompartmentach hematopoetycznych. Śledziona, migdałki, grasica, szpik kostny i krążące populacje komórek odpornościowych stanowiły zatem kanoniczne tkanki do analizy CB2.
To ukierunkowanie na układ odpornościowy ukształtowało wczesną koncepcję rozwoju leków, zgodnie z którą agoniści selektywni wobec CB2 mieli zapewniać korzyści przeciwzapalne lub przeciwbólowe przy jednoczesnym unikaniu efektów odurzających związanych z silną aktywacją CB1 w mózgu. Była to rozsądna hipoteza, ale tylko częściowo kompletna. CB2 jest GPCR sprzężonym z Gi/o i, podobnie jak CB1, nie działa wyłącznie jako przełącznik „włącz/wyłącz”. Zależnie od ligandu, konformacji receptora i kontekstu komórkowego CB2 może zmniejszać aktywność cyklazy adenylanowej, wpływać na szlaki MAPK, pośrednio modyfikować sprzężenie z kanałami jonowymi, rekrutować beta-arrestyny oraz ulegać desensytyzacji lub internalizacji. Zatem nawet w obwodowych tkankach odpornościowych rzeczywiste pytanie nie brzmi tylko, czy CB2 jest obecny, lecz które komórki go eksprymują, na jakim poziomie, pod wpływem jakiego bodźca i z jakim dalszym ukierunkowaniem sygnalizacji.
Ta złożoność jest jednym z powodów, dla których ligandy wyglądające podobnie mogą zachowywać się odmiennie. Przegląd z 2026 roku opublikowany w Frontiers in Chemical Biology dotyczący struktury receptora cannabinoidowego argumentuje, że selektywność wobec „CB1 i CB2” jest kształtowana przez różnice strukturalne na poziomie receptora, które zmieniają wiązanie ligandu, skuteczność sygnalizacji oraz regulację receptora. Badanie indeksowane w PubMed z lat 2025/2026 dotyczące selektywności podtypowej poszło dalej, pokazując, że selektywność endocannabinoidów jest dynamiczna i związana ze stanami konformacyjnymi, a nie ze sztywnym modelem klucz-zamek. Ma to znaczenie dla mapowania tkanek, ponieważ endogenny ligand taki jak 2-AG lub anandamid, phytocannabinoid taki jak THC oraz syntetyczny agonista preferujący CB2 mogą napotykać tę samą populację receptorów, a mimo to stabilizować odmienne wyjścia sygnałowe.
Starsza mapa CB2 skoncentrowana na układzie odpornościowym nie była zatem błędna. Była niepełna. CB2 nadal najlepiej opisywać jako receptor o silnych korzeniach w układzie odpornościowym. Ale korzenie nie stanowią całego organizmu.
| Kontekst | Jak opisano CB2 | Punkt interpretacyjny |
|---|---|---|
| Zdrowa baza mózgowa | Często niski lub bliski granicom wykrywalności w wielu regionach | Niski sygnał podstawowy nie oznacza braku znaczenia |
| Aktywowany mikroglej | Staje się bardziej wykrywalny po urazie lub zapaleniu | Wspiera znaczenie dla OUN poprzez funkcje podobne do immunologicznych |
| Astrocyty / śródbłonek / komórki naciekające | Opisywane w niektórych kontekstach chorobowych | Lokalizacja zależy od metody i modelu |
| Szeroka konstytutywna ekspresja neuronalna | Wymaga mocniejszych dowodów | Artykuł traktuje to twierdzenie ostrożnie |
CB2 w mikrogleju, neurozapaleniu i stanach pourazowych
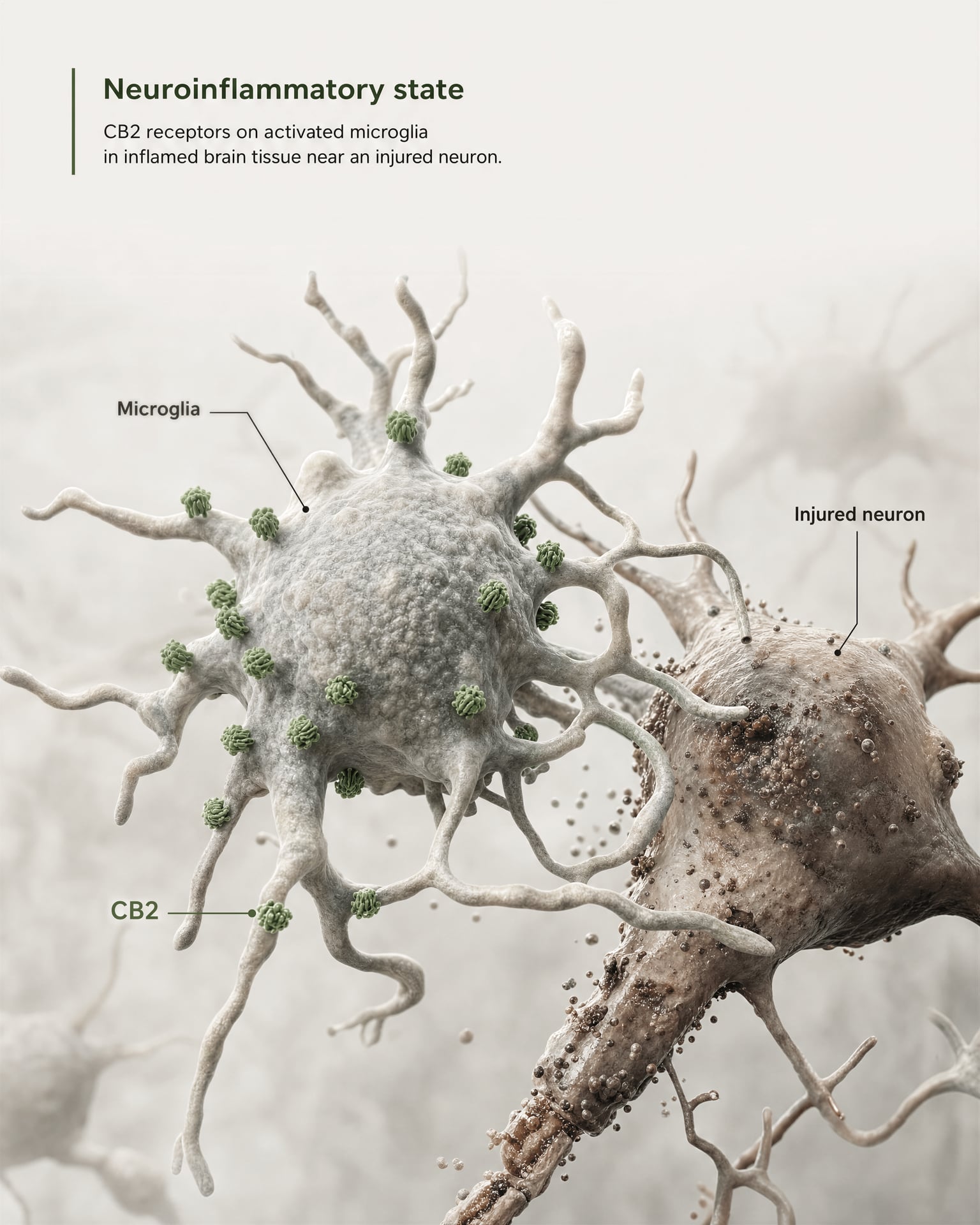
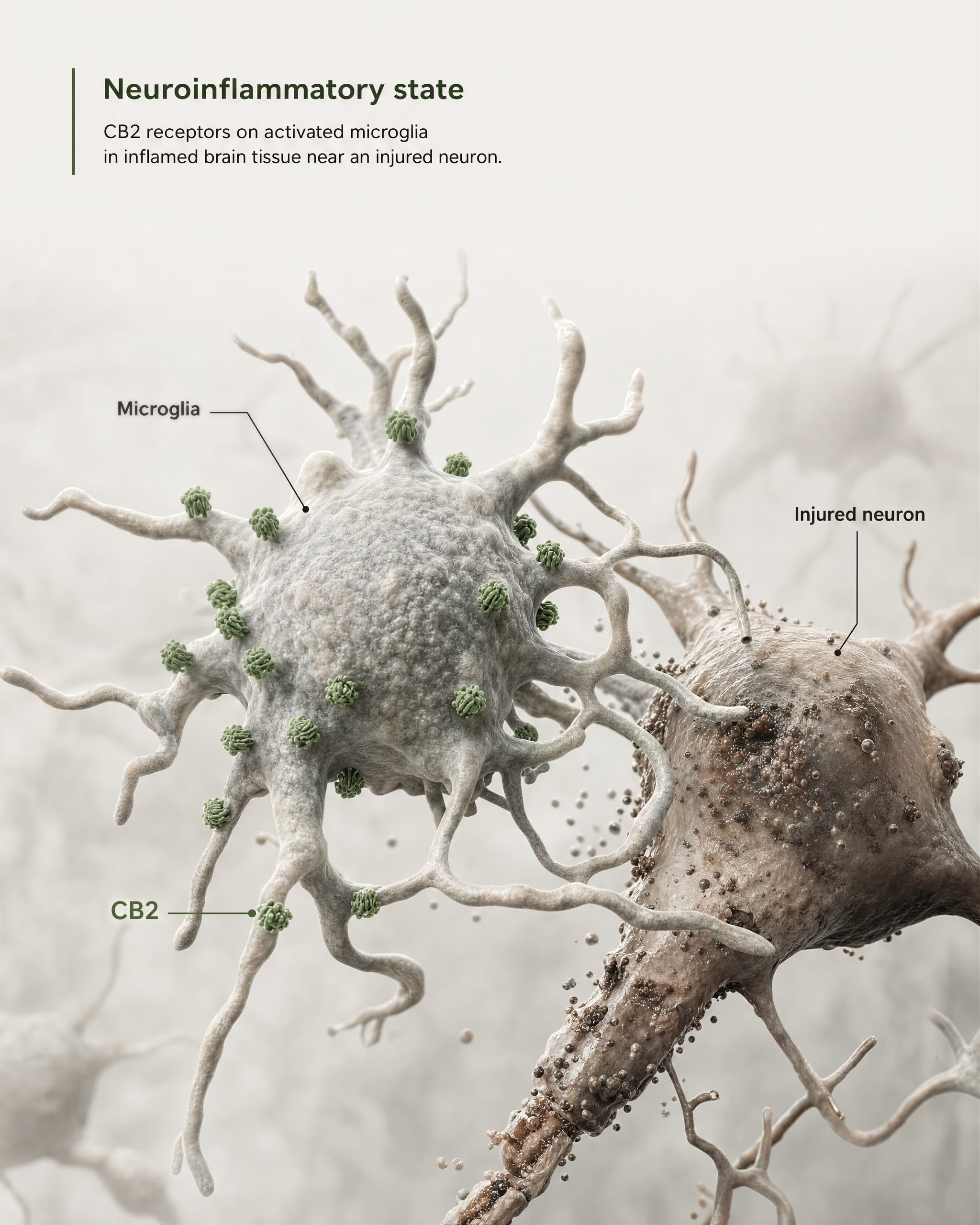
Najsilniejszy argument za znaczeniem ośrodkowym nie polega na twierdzeniu, że CB2 jest szeroko obecny w zdrowych neuronach przodomózgowia. Wynika on z mikrogleju i biologii chorób.
{kind=link}
Mikroglej to rezydentne komórki odpornościowe OUN i właśnie one wyznaczają granicę, na której stary model „obwodowego receptora układu odpornościowego” zaczyna zawodzić. Jeśli receptor jest eksprymowany w mózgowym własnym systemie nadzoru immunologicznego i odpowiedzi zapalnej, nazywanie go wyłącznie obwodowym staje się nieprecyzyjne. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience stawia tę kwestię wprost: sygnalizacja CB2 zyskała uwagę w zaburzeniach OUN, ponieważ wiąże się z mechanizmami neurozapalnymi i neurodegeneracyjnymi. Dlatego CB2 pojawia się dziś w dyskusjach o chorobie Alzheimera, chorobie Parkinsona, stwardnieniu rozsianym, urazowym uszkodzeniu mózgu, udarze, bólu neuropatycznym oraz niektórych zaburzeniach psychiatrycznych, w których sygnalizacja zapalna stanowi część patofizjologii.
Kluczowe jest określenie indukowana lub zwiększona ekspresja. W wielu zdrowych regionach mózgu bazalna ekspresja CB2 jest niska, czasem bliska granicom starszych metod detekcji. Jednak po urazie, infekcji, przewlekłym zapaleniu lub neurodegeneracji sygnał CB2 często staje się bardziej wykrywalny, zwłaszcza w aktywowanym mikrogleju, a w niektórych badaniach także w astrocytach, naciekających komórkach odpornościowych, kompartmentach śródbłonkowych lub ograniczonych podzbiorach neuronalnych. To zupełnie inna reguła rozmieszczenia niż ta zwykle stosowana wobec CB1. CB1 jest często konstytutywnie obfity w określonych obwodach neuronalnych. CB2 częściej interpretuje się jako receptor, którego znaczenie w OUN ujawnia się pod wpływem stresu, patologii lub aktywacji zapalnej.
To rozróżnienie ma praktyczne konsekwencje. Lek ukierunkowany na CB2 może wykazywać niewielki efekt w zdrowej tkance, gdzie gęstość receptora jest niska, a mimo to wykazywać mierzalną aktywność w tkance chorej, gdzie ekspresja wzrosła, a sieci sygnałowe uległy zmianie. Ta indukowalność jest jednym z powodów, dla których wyniki przedkliniczne były jednocześnie obiecujące i trudne do przełożenia na praktykę kliniczną. Liczy się czas. Liczy się stadium choroby. Liczy się skład komórkowy. Mikrośrodowisko mikrogleju po urazie nie jest farmakologicznie równoważne ze stymulowanym fragmentem mózgu.
Problemy interpretacyjne nie są błahe. CB2 ma długą historię zastrzeżeń dotyczących swoistości przeciwciał, wykrywania transkryptów o niskiej obfitości, różnic międzygatunkowych oraz niespójnych twierdzeń lokalizacyjnych w różnych metodach. Część wczesnych doniesień prawdopodobnie przeceniała neuronalny CB2, ponieważ dostępne narzędzia były słabe. Dlatego staranne badania opierają się obecnie na zbieżnych liniach dowodowych — transkryptomice pojedynczych komórek, hybrydyzacji in situ, zwalidowanych reporterach genetycznych, kontrolach knockout, danych proteomicznych tam, gdzie to możliwe, oraz porównaniach zależnych od stanu — zamiast na jednym wyniku barwienia. Jeśli jedno badanie wykazuje CB2 w neuronach w warunkach bazalnych, a inne nie potrafi go wykryć, rozbieżność może wynikać z rzeczywistych różnic regionalnych, stanu chorobowego, gatunku, wieku albo po prostu z ograniczeń testu.
CB2 ma istotne znaczenie dla OUN, zwłaszcza w kontekstach glejowych i zapalnych związanych z uszkodzeniem.Limited evidence
Najbardziej trafne stanowisko brzmi więc ostrożnie, ale jasno: CB2 ma realne znaczenie dla OUN, głównie poprzez funkcje glejowe i podobne do immunologicznych, a znaczenie to wzrasta podczas neurozapalania i urazu. Twierdzenia o szerokiej konstytutywnej ekspresji neuronalnej CB2 w prawidłowym mózgu wymagają mocniejszych dowodów niż twierdzenia o mikroglejowym i patologią związanym CB2.
Jak ostatnie 3 lata zmieniły rozmowę o CB2
Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience wyraźnie określa najnowszą literaturę jako „aktualizację z ostatnich 3 lat”, a to sformułowanie dobrze oddaje rzeczywistą zmianę. Dyskusja przesunęła się od sporu o to, czy CB2 „w ogóle jest w mózgu”, do pytań, gdzie, kiedy i w jakich stanach chorobowych jego sygnalizacja staje się terapeutycznie użyteczna.
Do tej zmiany przyczyniły się trzy rozwinięcia. Po pierwsze, ulepszyły się metody o wysokiej rozdzielczości komórkowej. Dane z RNA-seq pojedynczych komórek i pojedynczych jąder, lepsze mapowanie przestrzenne oraz bardziej rygorystyczne standardy walidacji zmniejszyły ryzyko, że niska lub indukowalna ekspresja zostanie odrzucona tylko dlatego, że starsze testy nie miały wystarczającej czułości. Po drugie, neurozapalanie stało się centralne dla wielu modeli chorób mózgu. Gdy choroby analizowano przez mechanizmy immunologiczne i glejowe, a nie wyłącznie przez pryzmat neuronów, CB2 stał się znacznie trudniejszy do zignorowania. Po trzecie, farmakologia receptorów dojrzała. Dziedzina myśli dziś bardziej w kategoriach skuteczności, biasu sygnałowego, transportu receptorów i odpowiedzi zależnych od kontekstu niż prostego zajęcia receptora.
Ta szersza perspektywa GPCR jest widoczna nawet poza literaturą dotyczącą CB2. Artykuł z 2025 roku w American Journal of Psychiatry o stronniczej sygnalizacji CB1 i schizofrenii argumentuje, że farmakologię cannabinoidową należy rozumieć przez pryzmat biasu sygnałowego, a nie surowej aktywacji receptora. Schizofrenia dotyka około 24 miliony ludzi na świecie, według WHO, więc nie jest to poboczna kwestia akademicka. Taka sama logika dotyczy CB2. Ligand „selektywny wobec CB2” na papierze może nadal wywoływać odmienne efekty zależnie od tego, czy preferencyjnie uruchamia sygnalizację białek G, rekrutację beta-arrestyn, internalizację receptora czy przeciwzapalne programy transkrypcyjne w aktywowanym mikrogleju.
Nowsza perspektywa systemowa to potwierdza. Badanie analizy sieciowej z 2025/2026 roku indeksowane w PubMed wskazało CB1 i CB2 jako silnie wpływowe węzły w systemie endocannabinoidowym i powiązało sygnalizację receptorową ze szlakami metabolicznymi, zamiast oddzielać receptory od reszty biologii komórki. Zgadza się to z danymi o CB2 w OUN: rozmieszczenie nie jest stałym wpisem atlasowym. Jest częścią adaptacyjnej sieci sygnałowej.
Wniosek jest prosty. CB2 nadal należy przedstawiać jako receptor cannabinoidowy wzbogacony w układzie odpornościowym. Jednak zatrzymanie się na tym obrazie daje dziś fałszywy obraz. W mózgu CB2 najlepiej rozumieć jako receptor o niskiej ekspresji bazalnej, indukowalny i związany z chorobą, którego znaczenie najwyraźniej ujawnia się w mikrogleju i stanach neurozapalnych — a jego wykrycie nadal w dużym stopniu zależy od metody, modelu i czasu.
Jak sygnalizują receptory cannabinoidowe: sprzężenie Gi/o, drugie przekaźniki i efekty synaptyczne
Farmakologia receptorów cannabinoidowych zaczyna się od prostego stwierdzenia, które bardzo szybko staje się złożone: CB1 i CB2 są receptorami sprzężonymi z białkiem G klasy A i oba najczęściej przekazują sygnał przez białka Gi/o. Ten podstawowy fakt, ustalony w pionierskich badaniach nad receptorami prowadzonych przez Allyn Howlett i innych, pozostaje aktualny. Zmieniło się natomiast rozumienie tego, co sprzężenie z Gi/o rzeczywiście oznacza w komórkach. Nie oznacza ono jednego efektu downstream. Oznacza zestaw możliwych efektów, których skład zależy od ligandu, gęstości receptora, stanu fosforylacji, środowiska błonowego, typu komórki i czasu.
To rozróżnienie ma znaczenie, ponieważ około 200 milionów ludzi używało cannabis w 2019 roku, czyli 4% globalnej populacji w wieku 15–64 lata według World Health Organization, podczas gdy FDA podaje, że w 2025 roku dopuszczono do obrotu jeden produkt leczniczy pochodzenia cannabis oraz trzy produkty lecznicze związane z cannabis. Sygnalizacja receptorowa nie jest kwestią uboczną. To mechanizm, który odróżnia użyteczny lek przeciwdrgawkowy od sedacji, nieudany lek pobudzający apetyt od psychiatrycznych działań niepożądanych oraz ligand selektywny w warunkach laboratoryjnych od związku rozczarowującego klinicznie.
Kanoniczna sygnalizacja GPCR w CB1 i CB2
Kanoniczna sekwencja sygnalizacji CB1/CB2
- Wiązanie ligandu Agonista stabilizuje aktywną konformację receptora.
- Aktywacja białka G Receptor promuje wymianę GDP na GTP w Gi/o.
- Rozdzielenie podjednostek Galpha i Gbeta-gamma regulują efektory dalszego szlaku.
- Zmiana przekaźnika wtórnego Aktywność cyklazy adenylanowej spada, a cAMP maleje.
- Efekt komórkowy Zmieniają się kanały jonowe, uwalnianie neuroprzekaźników, kinazy i regulacja genów.
W schemacie kanonicznym związanie agonisty stabilizuje aktywną konformację receptora, receptor działa jako czynnik wymiany nukleotydu guaninowego dla Gi/o, podjednostka Gαi/o wymienia GDP na GTP, a komponenty Gα i Gβγ następnie regulują efektory downstream. Dla CB1 i CB2 klasycznym odczytem jest hamowanie cyklazy adenylanowej i spadek wewnątrzkomórkowego cyklicznego AMP. To odkrycie stało się jednym z najwcześniejszych biochemicznych podpisów wykorzystywanych do definiowania aktywności receptora cannabinoidowego.
Jednak „kanoniczny” nie powinien być odczytywany jako „jednolity”. CB1 wykazuje wysoką aktywność konstytutywną w kilku systemach ekspresyjnych, co oznacza, że receptor może sygnalizować mierzalnie nawet bez obecności agonisty. Ta właściwość pomaga wyjaśnić, dlaczego antagoniści odwrotni, tacy jak rimonabant, nie tylko blokowali endogenny ton cannabinoidowy, lecz także obniżali sygnalizację poniżej poziomu wyjściowego i wywoływali wyraźne ośrodkowe działania niepożądane. CB2 również sprzęga się z Gi/o, jednak sposób, w jaki ligandy stabilizują stany aktywne, różni się od CB1. Prace strukturalne omówione w Frontiers in Chemical Biology w 2026 roku podkreśliły, że selektywność między „CB1 i CB2” wynika z różnic na poziomie receptora, które wpływają nie tylko na wiązanie, ale także na skuteczność i regulację. Badanie indeksowane w PubMed z 2025/2026 roku dotyczące selektywności podtypowej poszło dalej, argumentując, że selektywność endocannabinoidów ma charakter dynamiczny i jest kształtowana przez zachowanie konformacyjne, a nie przez stały model klucz–zamek.
To jeden z powodów, dla których phytocannabinoidy, endocannabinoidami i ligandy syntetyczne nigdy nie powinny być traktowane jako wymienne. Anandamid, zidentyfikowany przez Raphaela Mechoulama i Lumíra Hanuša, oraz 2-arachidonyloglicerol są endogennymi ligandami wytwarzanymi „na żądanie” i szybko inaktywowanymi. Δ9-THC jest pochodzącym z rośliny częściowym agonistą o kinetyce i skuteczności odmiennych od tych endocannabinoidów. Syntetyczne agoniści, takie jak CP55,940, WIN55,212-2 czy HU-210, często wywołują silniejszą aktywację receptora i mogą rekrutować szlaki sygnałowe w różnym stopniu. Niektóre ligandy preferują sygnalizację przez białko G kosztem rekrutacji β-arrestyny; inne nie. Artykuł w American Journal of Psychiatry z 2025 roku odniósł się do tego bezpośrednio w przypadku CB1, argumentując, że bias sygnałowy jest prawdopodobną strategią terapeutyczną w schizofrenii, zaburzeniu dotykającym około 24 milionów ludzi na świecie.
CB2 dodaje kolejną korektę do starszych uproszczeń. Nadal jest wzbogacony w wielu populacjach komórek układu odpornościowego, ale przegląd z 2026 roku w Frontiers in Behavioral Neuroscience opisał „aktualizację z ostatnich 3 lat”, w której sygnalizacja CB2 zyskała uwagę w zaburzeniach ośrodkowego układu nerwowego związanych z neurozapaleniem i neurodegeneracją. Zatem jeszcze zanim pojawi się zagadnienie agonizmu biasowanego, dawny podział na jeden „receptor mózgowy” i jeden „receptor immunologiczny” już na poziomie kontekstu sygnałowego okazuje się niewystarczający.
Efekty na cAMP, kanały jonowe i uwalnianie neurotransmiterów
Najlepiej znanym efektem drugiego przekaźnika w obu receptorach jest hamowanie powstawania cAMP poprzez inhibicję cyklazy adenylanowej. Niższe cAMP często oznacza mniejszą aktywność kinazy białkowej A, zmienioną fosforylację docelowych białek i wolniejsze zmiany ekspresji genów poprzez szlaki takie jak CREB. W neuronach jednak szybsze efekty są często ważniejsze niż wolniejsze.
CB1 jest silnie usytuowany do kontroli presynaptycznej. W wielu obwodach mózgowych znajduje się na zakończeniach aksonalnych, gdzie aktywacja receptora zmniejsza prawdopodobieństwo uwolnienia neurotransmitera. Dzieje się to poprzez połączenie hamowania kanałów wapniowych bramkowanych napięciem pośredniczonego przez Gβγ oraz aktywację przewodnictwa potasowego prostującego do wewnątrz lub innych prądów potasowych, które obniżają pobudliwość zakończenia. Mniejszy napływ wapnia oznacza mniej fuzji pęcherzyków. Skutkiem jest mniejsze uwalnianie neurotransmitera do szczeliny synaptycznej.
To jest podstawowy mechanizm krótkozasięgowej, retrogradowej sygnalizacji endocannabinoidowej. Neuron postsynaptyczny ulega aktywacji, syntetyzuje endocannabinoidy „na żądanie” z lipidowych prekursorów błonowych i wysyła je wstecz przez synapsę, aby aktywować presynaptyczne receptory CB1. Zakończenie presynaptyczne następnie uwalnia mniej neurotransmitera. Jest to hamulec zwrotny. W synapsach pobudzających może to tłumić uwalnianie glutaminianu; w synapsach hamujących może tłumić uwalnianie GABA. Kierunek wyjścia obwodu zależy od tego, które zakończenie eksprymuje CB1. Ten sam receptor, przeciwstawny skutek sieciowy.
| Termin | Co jest hamowane | Opisany mechanizm |
|---|---|---|
| DSI | Hamowanie | Aktywność postsynaptyczna uwalnia endokannabinoidy, które aktywują presynaptyczny CB1 i zmniejszają uwalnianie GABA |
| DSE | Pobudzenie | Aktywność postsynaptyczna uwalnia endokannabinoidy, które aktywują presynaptyczny CB1 i zmniejszają uwalnianie glutaminianu |
DSI i DSE Krótkotrwałe formy synaptycznej plastyczności pośredniczonej przez endokannabinoidy, w których depolaryzacja postsynaptyczna hamuje transmisję hamującą (DSI) lub pobudzającą (DSE) poprzez aktywację presynaptycznego CB1.
Klasyczne terminy fizjologiczne oddają to trafnie: depolaryzacyjnie indukowane hamowanie hamowania, DSI, oraz depolaryzacyjnie indukowane hamowanie pobudzenia, DSE. Oba są krótkotrwałymi formami plastyczności synaptycznej wywołanymi uwalnianiem endocannabinoidów i presynaptyczną aktywacją CB1. Występują także dłużej utrzymujące się efekty, w tym endocannabinoid-mediated long-term depression w niektórych synapsach. Zjawiska te mają znaczenie, ponieważ łączą biochemię receptora z zachowaniem: przetwarzanie bólu, wygaszanie lęku, uczenie nawyków, apetyt, kontrola motoryczna i próg drgawkowy zależą od tego dostrajania prawdopodobieństwa uwalniania.
Szczegóły nie są błahe. Częściowy agonista, taki jak Δ9-THC, może nie odtwarzać pełnego wzorca wywoływanego przez krótkotrwały endogenny wyrzut 2-AG. Także syntetyczny pełny agonista nie musi zachowywać fizjologicznego czasu działania. Znaczenie ma dawka. Znaczenie ma również rezerwa receptorowa. W synapsie o dużej gęstości CB1 nawet częściowy agonista może wywołać duży efekt na uwalnianie neurotransmitera. W innej tkance ten sam ligand może wyglądać na słaby.
CB2 ma mniej ustaloną bezpośrednią fizjologię synaptyczną niż CB1, ale również obniża cAMP i może regulować sygnalizację wapniową, szlaki kinaz oraz uwalnianie mediatorów zapalnych w komórkach odpornościowych i glejowych. To czyni CB2 istotnym dla komunikacji neuron–glej, zwłaszcza w stanach chorobowych, w których zmienia się ekspresja receptora. Niedawny artykuł oparty na analizie sieci i indeksowany w PubMed w latach 2025/2026 traktował CB1 i CB2 jako wpływowe węzły w szerszej sygnalizacji endocannabinoidowej i metabolicznej, co jest lepszym ujęciem niż przedstawianie ich jako izolowanych przełączników.
Desensytyzacja, internalizacja i regulacja receptora
Jak receptory dostosowują się do powtarzanej ekspozycji na agonistę
- Fosforylacja Wewnętrzne regiony receptora są modyfikowane przez kinazy GPCR i inne kinazy.
- Rekrutacja beta-arrestyn Arrestyny rozprzęgają receptory od białek G i mogą inicjować dodatkową sygnalizację.
- Desensytyzacja Receptor staje się mniej reaktywny.
- Internalizacja Receptory są wciągane do szlaków endocytozy.
- Los po wychwycie Receptory mogą być zawracane na powierzchnię albo degradowane.
Żaden receptor nie może być aktywowany w sposób ciągły bez konsekwencji. W przypadku CB1 i CB2 długotrwała lub powtarzana ekspozycja na agonistę zwykle prowadzi do fosforylacji wewnątrzkomórkowych regionów receptora przez kinazy GPCR i inne kinazy, rekrutacji β-arrestyn, odsprzęgnięcia od białek G, a następnie internalizacji przez szlaki endocytarne. Najpierw pojawia się desensytyzacja. Potem często następuje endocytoza. Następnie zachodzi recykling albo degradacja.
W przypadku CB1 ten cykl regulacyjny jest głównym powodem, dla którego ostre i przewlekłe efekty różnią się od siebie. Silne agoniści mogą wywoływać szybką desensytyzację w systemach komórkowych i mierzalną tolerancję in vivo. Znaczenie ma tu regulacja specyficzna dla regionu. Receptory CB1 nie ulegają desensytyzacji jednakowo we wszystkich populacjach neuronów, co pomaga wyjaśnić, dlaczego tolerancja rozwija się nierównomiernie w odniesieniu do różnych efektów cannabinoidowych. Odpowiedzi przeciwbólowe, hipotermia, zaburzenia pamięci i efekty motoryczne mogą zmieniać się w różnym tempie, ponieważ receptor jest regulowany odmiennie w poszczególnych obwodach.
β-arrestyny nie są jedynie przełącznikami wyłączającymi. Mogą one pełnić funkcję rusztowania dla własnych kaskad sygnałowych, w tym szlaków MAP kinazy, dlatego rekrutacja arrestyn stała się centralna dla agonizmu biasowanego. Ligand, który silnie hamuje cAMP, ale słabo rekrutuje β-arrestynę, może zachowywać się inaczej niż taki, który robi obie te rzeczy sprawnie. To nie jest już teoretyczny niuans; to strategia projektowania leków. Dyskusja z 2025 roku w American Journal of Psychiatry dotycząca biasu CB1 w schizofrenii odzwierciedla szerszą lekcję GPCR: unikanie niektórych gałęzi sygnalizacyjnych może zmniejszyć część ograniczeń, ale selektywność wobec szlaku nie gwarantuje sukcesu klinicznego.
Sama internalizacja także zależy od ligandu. Niektóre agoniści powodują rozległą endocytozę receptora; inne wywołują ograniczoną internalizację mimo aktywacji białka G. Modulatory allosteryczne dodatkowo komplikują obraz, ponieważ zmieniają sposób, w jaki ligandy ortosteryczne stabilizują stany receptora. W tym miejscu farmakologia strukturalna styka się z terapią. Przegląd strukturalny z 2026 roku jasno pokazał, że konformacja receptora kontroluje jednocześnie skuteczność sygnalizacji i regulację receptora, a nie są to odrębne zagadnienia.
To jest kluczowa lekcja sygnalizacyjna, którą należy zapamiętać. CB1 i CB2 nie są prostymi detektorami włącz/wyłącz dla cannabinoidów. Są regulowanymi węzłami, których wyjście zmienia się w skali od milisekund do dni. Każda poważna próba ich ukierunkowania, czy to w epilepsji, bólu, psychozie, czy chorobach zapalnych, musi uwzględniać sprzężenie Gi/o, drugie przekaźniki, kontrolę kanałów jonowych, lokalizację synaptyczną oraz fakt, że receptor będzie się adaptował do stymulacji.
Stronnicze sygnalizowanie: dlaczego jeden receptor może wywoływać różne skutki biologiczne
Stary obraz farmakologii cannabinoidów traktował receptor jak włącznik światła: agoniści go aktywują, antagoniści wyłączają, a wszystko inne wynika z tego, gdzie dany receptor jest eksprymowany. Taki obraz nie jest wystarczający dla CB1 ani CB2. Nie wyjaśnia, dlaczego dwa ligandy działające na tym samym receptorze mogą wywoływać wyraźnie odmienne efekty behawioralne, poznawcze, zapalne lub terapeutyczne. Nie wyjaśnia też, dlaczego badania nad receptorami cannabinoidowymi wielokrotnie prowadziły do związków, które in vitro wyglądały obiecująco, a następnie okazywały się rozczarowujące, nietolerowane albo klinicznie niejednoznaczne.
Ma to znaczenie daleko wykraczające poza akademicką teorię receptorów. World Health Organization oszacowała, że w 2019 roku 200 milionów ludzi używało cannabis, czyli około 4% globalnej populacji w wieku 15–64 lata. Schizophrenia dotyka około 24 milionów ludzi na świecie. Na tym tle farmakologia CB1 nie jest zagadnieniem niszowym. Znajduje się na styku zdrowia publicznego, psychiatrii i projektowania leków. U.S. FDA, według stanu na 2025 rok, zatwierdziła jeden lek pochodzący z cannabis oraz trzy produkty lecznicze związane z cannabis, co stanowi niewielką liczbę w porównaniu ze skalą zainteresowania klinicznego. Jednym z powodów, dla których postęp był wolniejszy, niż sugeruje debata publiczna, jest to, że sygnalizacja przez receptory cannabinoidowe nie polega na prostym zajęciu receptora. Chodzi o wybór szlaku.
Co oznacza stronnicze agonistyczne działanie w farmakologii GPCR
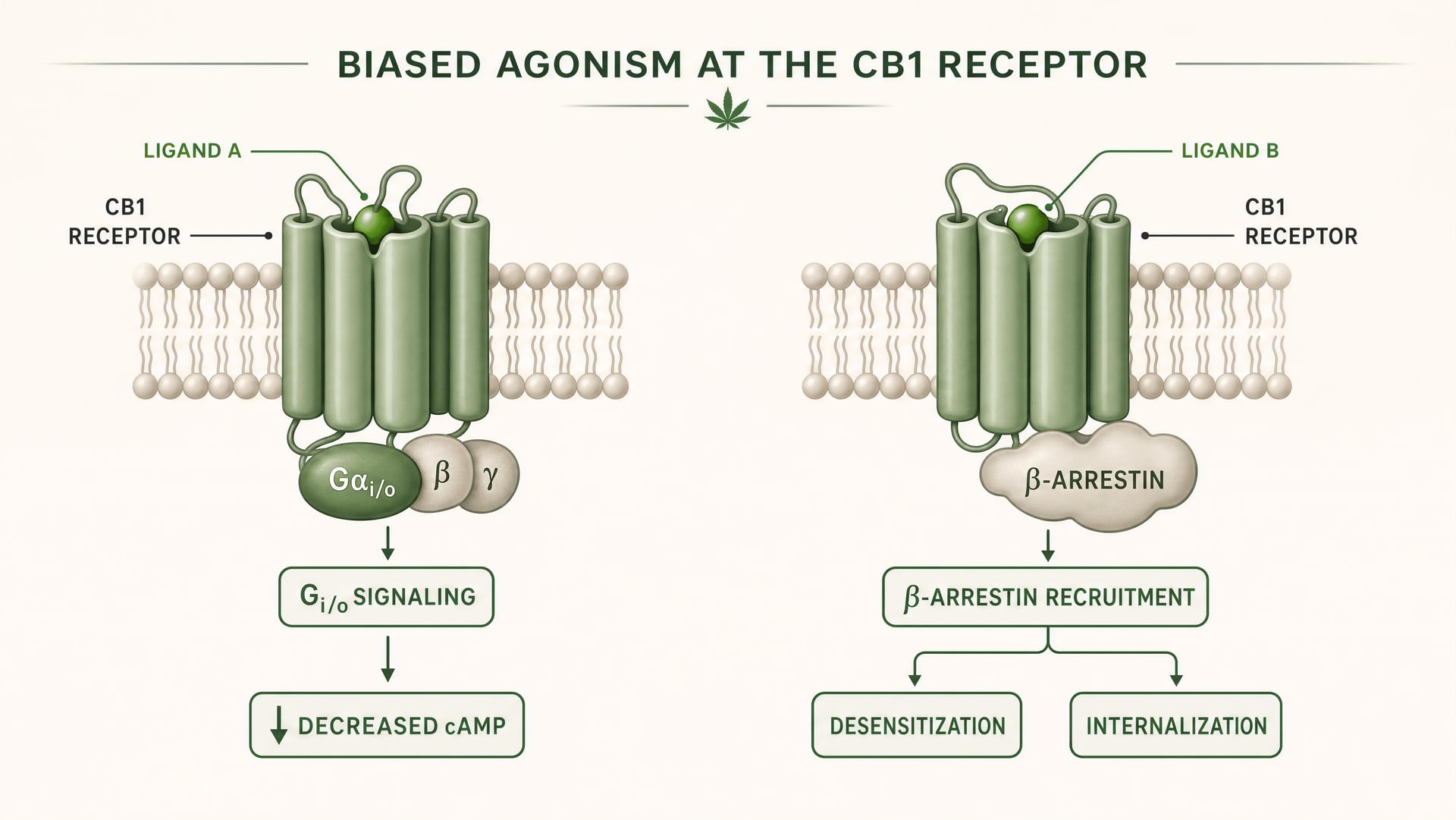
CB1 i CB2 są receptorami sprzężonymi z białkiem G klasy A. Fundamentalne prace Allyn Howlett ustanowiły CB1 jako receptor cannabinoidowy sprzężony z Gi/o, pomagając przenieść tę dziedzinę z niejasnej farmakologii do mechanizmów zdefiniowanych przez receptor. Jednak sprzężenie z Gi/o to dopiero początek. Po związaniu ligandu receptor może przyjąć więcej niż jedną aktywną konformację, a te konformacje nie sygnalizują identycznie. Niektóre konformacje receptora sprzyjają aktywacji białka G. Inne silniej rekrutują beta-arrestyny. Część stanów promuje fosforylację receptora, desensytyzację lub internalizację. Inne wywołują dłużej utrzymującą się sygnalizację z błony komórkowej albo z przedziałów endosomalnych.
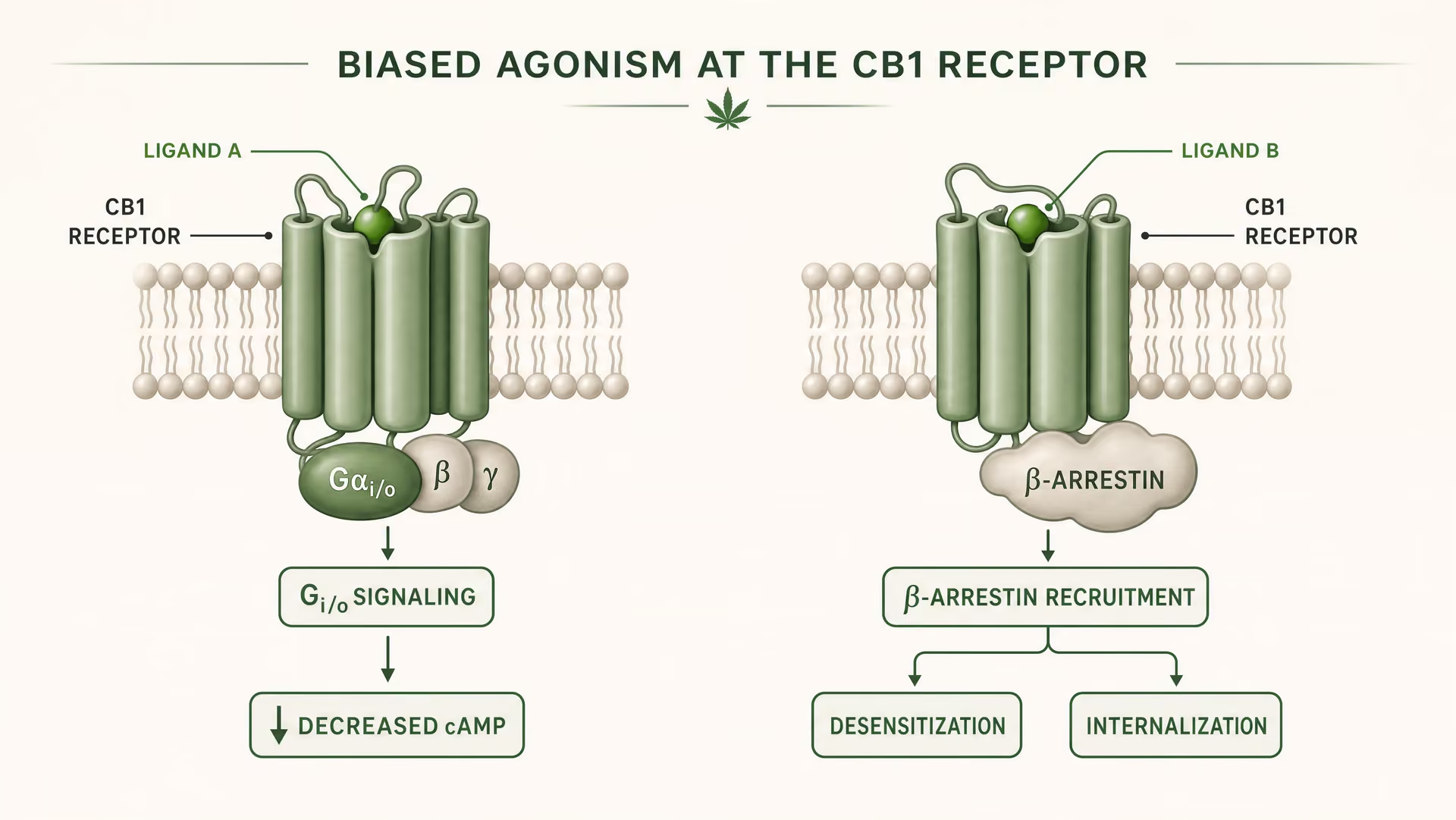{kind=link}
Agonizm biegunowany Właściwość ligandu, która faworyzuje niektóre dalsze szlaki sygnalizacyjne nad innymi w tym samym receptorze.
W prostych słowach stronnicze agonistyczne działanie oznacza, że różne ligandy stabilizują różne konformacje receptora, a te konformacje sprzyjają różnym szlakom dalszym. Receptor nie jest po prostu włączony albo wyłączony. Jest kierowany konformacyjnie.
W przypadku CB1 jest to szczególnie ważne, ponieważ receptor ten znajduje się w środowisku sygnalizacyjnym, które jest gęste, plastyczne i silnie zależne od typu komórki. W korowym zakończeniu glutaminergicznym ligand może zmniejszać uwalnianie neuroprzekaźnika poprzez hamowanie cyklazy adenylanowej zależne od Gi/o oraz modulację kanałów jonowych. W interneuronie GABAergicznym ten sam receptor może przesuwać lokalną równowagę obwodu neuronalnego w zupełnie innym kierunku. Jeśli ligand dodatkowo silnie promuje rekrutację beta-arrestyn, receptor może szybciej ulegać internalizacji, skracając jeden efekt i otwierając inny. Zmienia się czas. Zmienia się lokalizacja sygnału. Zmienia się odpowiedź fizjologiczna.
Nie jest to jedynie teoretyczne rozdrabnianie. Przegląd strukturalny z 2026 roku w Frontiers in Chemical Biology dotyczący receptorów cannabinoidowych jasno wskazuje, że selektywność ligandów wobec CB1 i CB2 zależy od różnic strukturalnych na poziomie receptora, które zmieniają wiązanie, skuteczność sygnałową i regulację receptora. Kluczowym słowem jest tu regulacja. Ligand może mieć podobne powinowactwo, a mimo to różnić się skutecznością, rekrutacją arrestyn, czasem przebywania przy receptorze lub skłonnością do wywoływania desensytyzacji. Badanie z 2025/2026 roku indeksowane w PubMed, dotyczące dynamicznego mechanizmu selektywności podtypów, rozwija tę samą ideę, argumentując, że selektywność wynika z dynamiki konformacyjnej, a nie ze statycznego modelu „zamek–klucz”. Endocannabinoids, phytocannabinoids i syntetyczne ligandy nie powinny być zatem wrzucane do jednego worka. Anandamide, odkryty przez Raphaela Mechoulama i Lumíra Hanuša, nie zachowuje się jak delta-9-tetrahydrocannabinol, a żaden z nich nie zachowuje się jak silnie zoptymalizowana syntetyczna sonda.
Stronnicze sygnalizowanie wyjaśnia również, dlaczego modulatory allosteryczne budzą tak duże zainteresowanie. Ligand allosteryczny może nie aktywować CB1 bezpośrednio tak jak agonista ortosteryczny, a mimo to może przekształcać preferencje sygnalizacyjne receptora, wzmacniając jeden szlak i tłumiąc inny. To otwiera drogę do precyzyjnej kontroli. W zasadzie.
Stronnicze sygnalizowanie CB1 jako kierunek badań nad schizofrenią
Artykuł z 2025 roku w American Journal of Psychiatry przedstawia najmocniejszy dotąd argument, że stronnicze sygnalizowanie CB1 nie jest jedynie pojęciem farmakologicznym, lecz możliwą strategią terapeutyczną w schizofrenii. Warto zwrócić na to uwagę, ponieważ badania nad schizofrenią zwykle traktowały cannabinoidy przez pryzmat epidemiologii, związku z ryzykiem albo ogólnych ostrzeżeń dotyczących psychozy. Artykuł w AJP zmienia perspektywę. Zadaje pytanie, czy problemem nie są „cannabinoidy” jako takie, ani nawet „aktywacja CB1” jako taka, lecz to, jakie stany sygnalizacyjne CB1 są uruchamiane, w których obwodach i przez jak długi czas.
To znacznie lepsze pytanie.
CB1 jest jednym z najliczniej występujących GPCR w mózgu, z dużą ekspresją w korze, hipokampie, zwojach podstawy i móżdżku, ale sama liczebność nie wyjaśnia efektów klinicznych. Schizofrenia obejmuje zaburzoną saliencję, poznanie, percepcję i koordynację sieciową w układach korowych i podkorowych. Receptor zdolny do modulowania aktywności obwodów glutaminergicznych, GABA-ergicznych i związanych z dopaminą jest zatem istotny z definicji. Artykuł w AJP argumentuje, że ligandy CB1 o stronniczym działaniu mogą oddzielić korzystne terapeutycznie efekty sieciowe od działań niepożądanych, takich jak pogorszenie funkcji poznawczych, lęk, dysforia czy reakcje psychotomimetyczne.
To ambitne twierdzenie, ale nie jest czystym spekulowaniem. Wynika z szerszej praktyki w dziedzinie GPCR, gdzie bias szlakowy już wcześniej zmienił sposób myślenia o lekach działających na receptory opioidowe, angiotensynowe i dopaminowe. Nadzieja translacyjna związana z CB1 polega na tym, że określone wyjścia sygnalizacyjne mogą poprawiać funkcję sieci korowej lub tłumić nieprawidłowe stany obwodowe bez odtwarzania pełnego profilu działań niepożądanych związanych z wysokoskuteczną agonistyką CB1.
Schizofrenia stanowi dobry przypadek testowy, ponieważ próg kliniczny jest wysoki. Kandydat na lek nie może jedynie zmieniać zachowania w badaniu na gryzoniach. Musi unikać nasilania psychozy, sedacji i zaburzeń poznawczych u osób już podatnych na te problemy. To natychmiast sprawia, że selektywność szlaku staje się czymś więcej niż preferencją chemii medycznej. Staje się wymogiem bezpieczeństwa.
Ujęcie z AJP pomaga też skorygować częste uproszczenie w dyskusjach o cannabis. Delta-9-THC jest phytocannabinoid z częściową aktywnością agonistyczną wobec CB1, ale jego działanie zależy od dawki, czasu, rezerwy receptorowej, lokalnego napięcia endocannabinoid i zaangażowania szlaków w różnych populacjach neuronów. Syntetyczny ligand CB1 zaprojektowany tak, by preferował jedną drogę wewnątrzkomórkową, może wyglądać zupełnie inaczej niż THC, nawet jeśli oba „działają na CB1”. Działa to też w drugą stronę: dwa związki, które poprawiają ten sam przedkliniczny punkt końcowy związany ze schizofrenią, mogą gwałtownie różnić się wpływem na poznanie lub afekt, jeśli jeden silnie uruchamia sygnalizację zależną od arrestyn, a drugi nie. Sama tożsamość receptora nie pozwala przewidzieć całego fenotypu.
Dlaczego selektywność szlaku ma znaczenie dla bezpieczeństwa i skuteczności
Selektywność szlaku ma znaczenie, ponieważ skuteczność nie jest jednowymiarowa. Lek cannabinoidowy może być silny, a mimo to klinicznie słaby. Może być selektywny wobec CB1 i nadal zawodzić. Może omijać CB2, a mimo to wywoływać niepożądane efekty immunologiczne lub metaboliczne poprzez sprzężenia sieciowe. Integracyjna analiza sieciowa z 2025/2026 roku, indeksowana w PubMed, zidentyfikowała CB1 i CB2 jako wysoce wpływowe węzły w systemie endocannabinoid i odwzorowała ich sygnalizację na szlaki metaboliczne. Taka perspektywa systemowa jest niezbędna. Receptory nie działają w izolacji, a stronniczość szlaku w jednym węźle może rozchodzić się na szersze programy fizjologiczne.
W przypadku CB1 obawy dotyczące bezpieczeństwa są oczywiste. Silna centralna aktywacja CB1 może powodować zaburzenia pamięci, zmienioną percepcję, lęk, tachykardię, a u osób podatnych także efekty związane z psychozą. Każdy program terapeutyczny ukierunkowany na ból, apetyt, nastrój, uzależnienie lub schizofrenię musi zmierzyć się z takim profilem ryzyka. Ligand, który zachowuje pożądany efekt synaptyczny zależny od Gi/o, a jednocześnie ogranicza desensytyzację napędzaną przez beta-arrestynę lub inne niekorzystne kaskady sygnałowe, mógłby teoretycznie poszerzyć okno terapeutyczne. Ale słowo „teoretycznie” ma znaczenie. Wiele programów z zakresu ligandów stronniczych w farmakologii GPCR pokazało, że bias mierzony w jednym układzie testowym nie zawsze przewiduje wynik in vivo. Tło komórkowe, gęstość receptorów, ekspresja efektorów i kinetyka mogą zmieniać pozorną stronniczość.
CB2 stanowi paralelne ostrzeżenie. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience opisuje aktualizację z ostatnich 3 lat, w której sygnalizacja CB2 zyskała uwagę w chorobach ośrodkowego układu nerwowego dzięki powiązaniom z mechanizmami neurozapalnymi i neurodegeneracyjnymi. Bezpośrednio podważa to dawną ideę, że CB2 jest dla mózgu nieistotny. Nawet tak nie jest, samo celowanie w CB2 nie gwarantuje użytecznego leku przeciwzapalnego. Rozmieszczenie jest bardziej stopniowane niż stary podział na mózg i ciało, a konsekwencje sygnalizacyjne nadal zależą od ligandu i kontekstu.
Praktyczny wniosek jest więc prosty: selektywność podtypu receptora jest konieczna, ale niewystarczająca. Selektywność szlaku może być różnicą między cannabinoidem, który wygląda terapeutycznie, tym, który działa odurzająco, i tym, który zawodzi w badaniach klinicznych, ponieważ nie potrafi oddzielić korzyści od działań niepożądanych. W przypadku CB1, zwłaszcza w psychiatrii, to właśnie ta różnica najprawdopodobniej zadecyduje, czy receptor pozostanie przestrogą, czy stanie się realnym celem lekowym.
Strukturalna biologia CB1 i CB2: jak kształt wpływa na selektywność
Biologia strukturalna zmieniła sposób, w jaki omawia się receptory cannabinoidowe. Stary skrót myślowy — CB1 wyjaśnia odurzenie, CB2 wyjaśnia stan zapalny — pomija fakt, że oba receptory są receptorami sprzężonymi z białkiem G klasy A, których zachowanie zależy od kształtu, ruchu, głębokości wiązania oraz partnerów sygnałowych dostępnych w danej komórce. Ma to znaczenie daleko wykraczające poza podstawową farmakologię. Szacuje się, że cannabis było używane przez około 200 milionów osób w 2019 roku, czyli przez 4% światowej populacji w wieku 15–64 lat, według WHO, jednak FDA nadal wskazuje tylko jeden produkt leczniczy pochodzenia cannabis oraz trzy produkty lecznicze związane z cannabis jako dopuszczone do obrotu w 2025 roku. Jedną z przyczyn tej luki jest struktura: trudno jest zaprojektować ligandy cannabinoidowe, które trafiają we właściwy receptor, we właściwy sposób, i przez odpowiednio długi czas.
Przegląd z 2026 roku opublikowany w Frontiers in Chemical Biology jasno to pokazuje. CB1 i CB2 różnią się nie tylko miejscem ekspresji. Różnią się architekturą kieszeni wiążących ligand, kształtem i elastycznością pętli zewnątrzkomórkowych, upakowaniem helis transbłonowych oraz stanami konformacyjnymi, które preferują po związaniu ligandu. Cechy te wpływają nie tylko na selektywność, lecz także na skuteczność, desensytyzację, internalizację i bias szlakowy.
Co badania strukturalne ujawniają o kieszeniach receptorów
Kieszeń ortosteryczna to główna jama wiążąca, w której endogenne ligandy, takie jak anandamid i 2-arachidonoiloglicerol, fitocannabinoidy, takie jak THC, oraz wiele ligandów syntetycznych nawiązują podstawowy kontakt. W CB1 i CB2 kieszeń ta znajduje się wewnątrz wiązki siedmiu helis transbłonowych, częściowo przykryta przez pętle zewnątrzkomórkowe, które mogą albo ułatwiać dostęp, albo go ograniczać.
Struktury kriomikroskopii elektronowej i rentgenowskie z ostatnich kilku lat pokazały, że receptory cannabinoidowe nie zachowują się jak sztywne zamki czekające na klucz. Lepiej rozumieć je jako ruchome cele o preferowanych kształtach. Przegląd z 2026 roku w Frontiers in Chemical Biology podkreśla, że kieszenie ortosteryczne CB1 i CB2 są na tyle podobne, by wiązać nakładające się klasy ligandów, a jednocześnie na tyle różne pod względem rozmiaru, tożsamości reszt i lokalnej elastyczności, by przesuwać powinowactwo i wynik sygnalizacyjny. Dlatego blisko spokrewnione związki mogą różnić się farmakologicznie. Niewielka zmiana wielkości podstawniku, polarności lub długości łańcucha może zmienić głębokość penetracji ligandu do kieszeni, helisy, które są przez niego odpychane, oraz to, czy receptor przyjmuje stan sprzyjający białkom G czy arrestynom.
CB1 okazał się szczególnie pouczający strukturalnie, ponieważ istnieje już wiele modeli o wysokiej rozdzielczości, zarówno w stanie nieaktywnym, jak i aktywnym. Powracającym motywem jest to, że jego kieszeń jest rozległa i hydrofobowa, co odpowiada lipofilowemu charakterowi wielu cannabinoidów. Pętla zewnątrzkomórkowa 2 oraz górne części kilku helis pomagają kształtować wejście. Helisy transbłonowe to siedem segmentów przebijających błonę, które tworzą rdzeń receptora; po związaniu ligandu mogą one przesuwać się względem siebie. Najważniejszy z farmakologicznego punktu widzenia ruch zachodzi zwykle po stronie wewnątrzkomórkowej, gdzie ruch helisy 6 na zewnątrz pomaga utworzyć miejsce dokowania dla białek Gi/o. To przesunięcie jest jednym z wyróżników aktywacji receptora.
CB2 ma ten sam ogólny fałd GPCR, ale przegląd Frontiers argumentuje, że różnice aminokwasowe specyficzne dla podtypu, wokół kieszeni i w regionach pętli, dają chemikom medycznym uchwyty do selektywności. Nie chodzi o to, że jedna kieszeń jest po prostu „mózgowa”, a druga „immunologiczna”. Chodzi o geometrię i energię. Odmienne reszty zmieniają kontur jamy, lokalne możliwości wiązań wodorowych, układanie aromatyczne i elastyczność kanałów dostępu, przez które ligandy wchodzą z błony.
Badanie indeksowane w PubMed z 2025/2026 roku dotyczące dynamicznego mechanizmu selektywności podtypowej poszło dalej, argumentując, że selektywność endocannabinoidów nie jest jedynie kwestią statycznego powinowactwa wiązania. Liczą się dynamika konformacyjna. Mówiąc prościej, receptor może próbować wielu kształtów przed i po związaniu ligandu, a niektóre ligandy stabilizują określony kształt lepiej niż inne. Pomaga to wyjaśnić, dlaczego lipidy endogenne, fitocannabinoidy i ligandy syntetyczne mogą wykazywać różne preferencje podtypowe, nawet gdy ich szkielety chemiczne na papierze wydają się podobne.
Czynniki determinujące selektywność ligandów między CB1 i CB2
Selektywność zaczyna się od chemii kontaktu, ale na niej się nie kończy. Przegląd w Frontiers in Chemical Biology ujmuje selektywność jako produkt strukturalnych różnic na poziomie receptora, które jednocześnie wpływają na wiązanie, skuteczność sygnałową i regulację. I to jest właściwe ujęcie. Ligand może być selektywny wobec CB2 w badaniu wiązania radioligandów, a mimo to utracić praktyczną przewagę, jeśli jednocześnie indukuje stany receptora powodujące szybką tolerancję lub słabe sygnalizowanie w komórkach istotnych dla choroby.
Kilka cech strukturalnych powraca wielokrotnie. Po pierwsze, skład aminokwasowy kieszeni ortosterycznej różni się na tyle między CB1 i CB2, by zmieniać sposób dopasowania grupy głowy ligandu, rdzenia i hydrofobowego ogona. Po drugie, pętle zewnątrzkomórkowe pomagają kształtować wejście i orientację. Po trzecie, górne i środkowe regiony helis transbłonowych mogą przesuwać receptor w kierunku nieco odmiennych zespołów stanów aktywnych. Stan konformacyjny to po prostu jeden z możliwych kształtów receptora w danym momencie. Różne ligandy nie tylko wiążą się z receptorem; stabilizują one podzbiór tych kształtów.
Dlatego selektywność podtypowa jest często umiarkowana w przypadku naturalnych cannabinoidów. THC na przykład oddziałuje z oboma receptorami. Anandamid i 2-AG również działają na oba, choć z zależnymi od kontekstu różnicami w mocy działania, skuteczności i metabolizmie. Ligandy syntetyczne były bardziej użyteczne do rozplątywania relacji struktura-selektywność, ponieważ chemicy mogą systematycznie modyfikować takie cechy jak długość łańcucha bocznego, ograniczenia pierścieniowe i polarne podstawnik. Nawet wtedy uzyskanie wyraźnego rozdzielenia jest trudne. CB1 i CB2 są na tyle homologiczne, że związek zaprojektowany pod jeden często zachowuje istotną aktywność wobec drugiego.
Ma to praktyczne konsekwencje. Twórcy leków od dawna poszukują agonistów selektywnych wobec CB2 z nadzieją na uzyskanie działania przeciwzapalnego lub przeciwbólowego bez silnych ośrodkowych działań niepożądanych zależnych od CB1. Czasem taka strategia działa farmakologicznie, ale nie jest to proste obejście problemu. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience podkreśla, że CB2 w ciągu „ostatnich 3 lat” przyciągnął uwagę w zaburzeniach ośrodkowego układu nerwowego, podważając uproszczone przekonanie, że CB2 jest nieistotny dla mózgu. Zatem nawet „obwodowy” ligand CB2 nie może być interpretowany w oparciu o przestarzałe mapy receptorów.
Dlaczego skuteczność i regulacja są również kwestiami strukturalnymi
Moc działania odpowiada na pytanie, ile ligandu potrzeba. Skuteczność odpowiada na pytanie, co ligand robi z receptorem po związaniu. Biologia strukturalna łączy te dwie kwestie, ale ich nie miesza. Dwa ligandy mogą zajmować tę samą kieszeń i wywoływać bardzo różne odpowiedzi, ponieważ stabilizują odmienne konformacje aktywne.
W przypadku CB1 ma to centralne znaczenie dla współczesnego myślenia terapeutycznego. Artykuł w American Journal of Psychiatry z 2025 roku argumentuje, że biasowane sygnalizowanie przez CB1 może być obiecującą strategią w schizofrenii, zaburzeniu dotykającym około 24 milionów osób na świecie. Agonizm z biasem oznacza, że ligand faworyzuje jeden szlak downstream kosztem innego, często sygnalizację Gi/o zamiast rekrutacji beta-arrestyny, lub odwrotnie. Strukturalnie taki bias wynika z tego, że ligand przestawia wewnątrzkomórkową powierzchnię receptora do kształtu lepiej pasującego do jednego partnera sygnałowego niż do innego. To nie jest abstrakcyjna idea. To cel chemii medycznej.
Beta-arrestyny mają znaczenie, ponieważ uczestniczą w desensytyzacji i internalizacji. Desensytyzacja oznacza, że receptor staje się mniej wrażliwy po powtarzającej się lub długotrwałej aktywacji. Internalizacja oznacza, że receptor zostaje wciągnięty z błony powierzchniowej do wnętrza komórki. Oba procesy są zależne od konformacji receptora. Niektóre ligandy silnie aktywują białka G, ale słabo rekrutują arrestyny; inne robią oba te rzeczy. Dobrze tolerowany lek działający na CB1 może potrzebować właśnie takiego rozdzielenia, a nie prostego blokowania lub pełnego agonizmu.
Dodatkową warstwę stanowi modulacja allosteryczna. Ligand allosteryczny wiąże się poza kieszenią ortosteryczną i zmienia sposób, w jaki receptor reaguje na inne ligandy. Strukturalnie jest to atrakcyjne, ponieważ może pozwolić na subtelniejszą kontrolę kształtu receptora niż intensywne pobudzenie głównej kieszeni przez agonistę o wysokiej skuteczności. W farmakologii cannabinoidowej mogłoby to oznaczać zachowanie korzystnej sygnalizacji przy jednoczesnym ograniczeniu działań niepożądanych związanych z nadmierną aktywacją CB1.
Wniosek ogólny jest jednoznaczny. Kształt receptora nie jest pobocznym szczegółem. To wyjaśnienie, dlaczego fitocannabinoidy, endocannabinoidy i związki syntetyczne mogą wywoływać odmienne profile kliniczne, mimo że celują w tę samą rodzinę receptorów. Ponieważ CB1 i CB2 zajmują wpływowe węzły w analizach sieciowych systemu endocannabinoid, jak wykazało badanie systemowe indeksowane w PubMed z 2025/2026 roku, błędy w selektywności lub skuteczności mogą rozprzestrzeniać się na wiele szlaków. Wgląd strukturalny nie gwarantuje sukcesu leku. Wyjaśnia jednak, dlaczego tak trudno jest dopasować do siebie selektywność, moc działania i tolerancję.
Endokannabinoidy i selektywność podtypowa: anandamid, 2-AG oraz dynamiczne zachowanie receptorów
Farmakologia endokannabinoidów jest często wyjaśniana za pomocą skrótu myślowego, który brzmi klarownie, ale wprowadza w błąd: anandamid i 2-arachidonoiloglicerol (2-AG) traktuje się tak, jakby były wbudowanymi przez organizm „standardowymi agonistami”, użytecznymi głównie jako punkty odniesienia dla tego, co zwykle robią CB1 i CB2. Taka rama pomija jednak sprawę zasadniczą. Nie są to stabilne, swobodnie krążące hormony, które napotykają receptory w jednolitych warunkach. Są to krótkotrwałe lipidowe mediatory wytwarzane na żądanie, w błonach komórkowych, przez swoiste dla danej komórki układy enzymatyczne, a następnie szybko rozkładane. Ich pozorna preferencja receptorowa może zmieniać się w zależności od składu błony, konformacji receptorów, lokalnej aktywności enzymów oraz czasu uwalniania.
Ma to znaczenie wykraczające poza akademicką teorię receptorów. Szacuje się, że w 2019 r. cannabis było używane przez około 200 milionów osób, czyli 4% światowej populacji w wieku 15–64 lat, zgodnie z WHO, a leki ukierunkowane na cannabinoid już są stosowane klinicznie: FDA podała w 2025 r., że zatwierdziła jeden produkt leczniczy pochodzący z cannabis oraz trzy produkty lecznicze związane z cannabis. Jeśli ligandy endogenne nie zachowują się jak proste podręcznikowe kontrole, to porównywanie fitokannabinoidów lub kandydatów na leki z nimi nie jest sprawą prostą.
Dlaczego ligandy endogenne nie są prostymi związkami referencyjnymi
Anandamid, zidentyfikowany w 1992 r. przez Devane’a, Hanusa, Breuera, Mechoulama i współpracowników, oraz 2-AG, uznany kilka lat później za kolejny główny endokannabinoid, różnią się nie tylko tabelami siły działania. Pochodzą z odmiennych szlaków biosyntezy, docierają do receptorów w różnych skalach czasowych i są usuwane przez różne enzymy. Anandamid powstaje głównie z N-arachidonoilo-fosfatydyloetanoloaminy z udziałem szlaków obejmujących NAPE-PLD, choć istnieją też alternatywne drogi. 2-AG jest wytwarzany przede wszystkim z diacyloglicerolu przez DAGLα i DAGLβ. Nie są to drobne biochemiczne przypisy; to one określają, gdzie i kiedy dochodzi do aktywacji receptorów.
Nawet starsze twierdzenie, że anandamid jest ligandem preferującym CB1, podczas gdy 2-AG jest pełnym agonistą zarówno CB1, jak i CB2, może okazać się zbyt uproszczone, gdy przechodzi się z układów oczyszczonych do żywej tkanki. Anandamid jest także substratem dla hydrolazy amidów kwasów tłuszczowych, czyli FAAH, i w pewnych warunkach może oddziaływać z celami innymi niż cannabinoid, takimi jak TRPV1. 2-AG występuje w tkance mózgowej na wielu pomiarach na znacznie wyższych poziomach bazalnych niż anandamid i jest hydrolizowany głównie przez lipazę monoacyloglicerolową, czyli MAGL, przy udziale także ABHD6 i ABHD12. Tak więc „sygnał wejściowy”, jaki każdy z ligandów dostarcza receptorowi, nie jest tylko kwestią powinowactwa czy skuteczności wobec CB1 i CB2. Zależy on od tego, czy ligand jest wytwarzany w synapsie, wewnątrz komórki odpornościowej, w pobliżu błony wewnątrzkomórkowej albo w przedziale bogatym w enzymy degradujące.
Prace Allyn Howlett z zakresu farmakologii receptorowej pomogły ustalić, że CB1 jest rzeczywistym GPCR, sprzężonym w dużej mierze z białkami Gi/o. Ta rama pozostaje niezbędna, ale nie wspiera już prostego modelu endogennego ligandu jako punktu odniesienia dla leków. CB1 i CB2 mogą sygnalizować poprzez hamowanie cyklazy adenylanowej, regulację kanałów jonowych, szlaki kinazy MAP, rekrutację beta-arrestyny, internalizację receptorów oraz desensytyzację. Ligand, który w jednym teście wygląda na „słabszy”, może wywoływać dłuższy sygnał w tkance, gdzie rozkład zachodzi wolno, albo może faworyzować jeden szlak sygnałowy kosztem innego. Artykuł z 2025 r. w American Journal of Psychiatry dotyczący biasowanego sygnalizowania CB1 podkreśla tę kwestię w kontekście choroby: w zaburzeniach takich jak schizofrenia, która dotyczy około 24 milionów osób na świecie, pytanie terapeutyczne nie brzmi jedynie, czy CB1 zostaje aktywowany, ale w jaki sposób.
Dynamiczne mechanizmy selektywności podtypowej
Niedawne badanie dotyczące selektywności podtypowej, indeksowane w PubMed (PMID: 41962866), jest ważne, ponieważ odchodzi od opowieści typu „zamek i klucz”. Jego zasadniczy przekaz brzmi, że selektywność endokannabinoidów może wynikać z mechanizmów dynamicznych. Innymi słowy, preferencji wobec podtypu receptora nie wyjaśnia wyłącznie stała różnica w sile wiązania ligandu do CB1 w porównaniu z CB2. Kompleks ligand-receptor próbuje wielu stanów konformacyjnych, a stany te różnią się między CB1 i CB2.
To odkrycie jest zgodne z przeglądem z 2026 r. w Frontiers in Chemical Biology, który wskazuje, że strukturalne różnice między CB1 i CB2 kształtują wiązanie, skuteczność sygnalizacji oraz regulację receptorów. Receptory te wykazują znaczną podobieństwo sekwencyjne, jednak niewielkie różnice w architekturze kieszeni ortosterycznej, pętlach zewnątrzkomórkowych, ruchach helis transbłonowych oraz powierzchniach sprzęgających po stronie wewnątrzkomórkowej mogą wpływać na to, co dzieje się po związaniu ligandu. Ligand może wchodzić do podobnych kieszeni w obu receptorach, ale stabilizować w jednym podtypie bardziej kompetentny sygnałowo stan albo faworyzować różne przejścia między stanami nieaktywnym, pośrednim i aktywnym.
W przypadku endokannabinoidów jest to szczególnie prawdopodobne, ponieważ są one elastycznymi lipidami, a nie sztywnymi rusztowaniami syntetycznymi. Elastyczność jest zmienną farmakologiczną. Anandamid może przyjmować wiele konformacji, a 2-AG jest równie dynamiczny w błonach. Pytanie brzmi więc nie tylko: „Czy się wiąże?”, lecz także: „Które stany receptora preferuje, jak długo stany te są zajęte i jakie białka są dostępne do dalszego sprzęgania?” Neuron bogaty w CB1 i komórka mikrogleju wykazująca ekspresję CB2 nie przedstawiają tego samego środowiska sygnałowego, nawet zanim uwzględni się różnice w gęstości receptorów.
Ta dynamiczna perspektywa osłabia także dawny pogląd, że ligandy endogenne wyznaczają jedną fizjologiczną bazę, względem której można klasyfikować THC, cannabidiol lub syntetyczne cannabinoids. THC jest fitokannabinoidem o własnym wzorcu skuteczności i zachowaniu kinetycznym. Endokannabinoidy są sygnałami wyzwalanymi zdarzeniowo, często fazowymi i lokalnymi. Syntetyczny agonista CB2 może wyglądać na selektywny w teście rekombinowanym, a mimo to dawać nieoczekiwane wyniki w tkance, jeśli różnią się rezerwa receptorowa, cholesterol błonowy, tworzenie heteromerów lub obsługa przez beta-arrestynę. Selektywność na papierze nie jest przeznaczeniem.
Jak lokalna synteza i degradacja kształtują oddziaływanie z receptorami
In vivo sygnalizacja endokannabinoidowa jest regulowana równie silnie przez enzymologię, jak przez farmakologię receptorową. W wielu synapsach ośrodkowego układu nerwowego 2-AG powstaje postsynaptycznie w odpowiedzi na wzrost stężenia wapnia lub aktywację receptorów sprzężonych z Gq/11, a następnie przemieszcza się retrogradowo do presynaptycznych receptorów CB1 i hamuje uwalnianie neuroprzekaźników. Jest to mechanizm czasowy mierzony w sekundach, a nie rozproszony ton tła. Sygnał kończy się, gdy MAGL i pokrewne hydrolazy usuwają 2-AG. Anandamid często zachowuje się inaczej, z mniejszą obfitością w tkankach, odmiennymi warunkami uwalniania oraz większą wrażliwością na rozkład kontrolowany przez FAAH.
Oznacza to, że zmiana aktywności enzymu może zmienić oddziaływanie z receptorami bez jakiejkolwiek zmiany samego receptora. Hamowanie FAAH zwiększa ton anandamidu, ale wynik fizjologiczny zależy od tego, gdzie anandamid jest wytwarzany, czy obecny jest również TRPV1 oraz czy receptory CB1 w danym mikroobszarze są zdesensytyzowane lub zinternalizowane. Hamowanie MAGL może znacząco podnieść poziom 2-AG, ale przewlekły wzrost może w niektórych układach prowadzić do desensytyzacji CB1. Więcej ligandu nie zawsze oznacza bardziej użyteczną sygnalizację.
Badanie systemowo-biologiczne indeksowane jako PMID: 42129940 wzmacnia ten szerszy obraz, umieszczając CB1 i CB2 wśród istotnych węzłów powiązanych ze szlakami metabolicznymi, a nie jako odizolowane przełączniki. To właściwa skala do rozumienia działania endokannabinoidów. Dostępność ligandu, dostęp do błony, konformacja receptora, sprzęganie z białkiem G, rekrutacja beta-arrestyny oraz degradacja wszystko to wzajemnie na siebie oddziałuje. Podobnie jak gradienty tkankowe. CB2 nie jest po prostu „poza mózgiem”, a CB1 nie jest wyłącznie „receptorem odpowiedzialnym za psychoaktywność”. Sygnalizacja endokannabinoidowa jest lokalna, zależna od warunków i kinetyczna. Każde ujęcie selektywności podtypowej, które ignoruje te cechy, błędnie odczyta zarówno fizjologię, jak i rozwój leków.
Układ endokannabinoidowy jako sieć, a nie diagram dwóch receptorów
Układ endokannabinoidowy ma niewielki sens, jeśli przedstawia się go jako dwa okręgi oznaczone CB1 i CB2 ze strzałkami skierowanymi na zewnątrz. Taki uproszczony schemat był pomocny we wczesnym okresie rozwoju tej dziedziny, zwłaszcza po pracach Allyn Howlett nad farmakologią receptorów i identyfikacji anandamidu przez Raphaela Mechoulama oraz Lumíra Hanuša, ale dziś bardziej zaciemnia obraz, niż go wyjaśnia. Stawka praktyczna jest duża. WHO szacuje, że w 2019 roku 200 milionów ludzi używało cannabis, czyli około 4% światowej populacji w wieku 15–64 lata, podczas gdy FDA podaje, że według stanu na 2025 rok zatwierdzono jeden produkt leczniczy pochodzenia cannabis oraz trzy produkty lecznicze związane z cannabis. System receptorowy obejmujący tak wiele ekspozycji i tak wiele twierdzeń terapeutycznych nie może być traktowany jako prosta para: receptor mózgowy plus receptor immunologiczny.
Podejście systemowe zaczyna się od podstawowego faktu: endokannabinoidy są lipidami wytwarzanymi na żądanie, szybko ulegającymi degradacji i osadzonymi w tej samej przestrzeni biochemicznej co mediatory zapalne, szlaki przebudowy błon i pętle sprzężenia zwrotnego synaptycznego. Anandamid i 2-arachidonyloglicerol nie są odosobnionymi „komunikatami”. Powstają z prekursorów błonowych, konkurują z innymi szlakami lipidowymi o substraty i działają w tkankach, w których gęstość receptorów, typ komórki, ekspresja enzymów oraz partnerzy sygnalizacyjni są różne. Dlatego ten sam ligand może wyglądać na anksjolityczny w jednym kontekście, odurzający w innym, przeciwzapalny w jeszcze innym, a klinicznie rozczarowujący w badaniu.
Co wnosi integracyjna analiza sieciowa
Badanie integracyjnej analizy sieciowej indeksowane w PubMed, opublikowane w 2025/2026 roku (PMID: 42129940), jest użyteczne, ponieważ przesuwa uwagę z pojedynczych celów na organizację całego systemu. Zamiast pytać wyłącznie, który ligand wiąże CB1 lub CB2, autorzy mapują układ endokannabinoidowy jako sieć interakcji obejmującą receptory, enzymy biosyntetyczne i degradacyjne, lipidowe intermedianty oraz powiązania na poziomie szlaków z zapaleniem, metabolizmem i sygnalizacją neuronalną. W takim ujęciu wpływ nie jest definiowany jedynie przez liczebność receptorów. Określa go łączność oraz sposób, w jaki zaburzenia rozprzestrzeniają się po sieci.
Ma to znaczenie dla interpretacji. Jeśli lek zmienia sygnalizację CB1, efekt downstream nie ogranicza się do hamowania cyklazy adenylanowej pośredniczonego przez Gi/o. Może on zmieniać uwalnianie neuroprzekaźników, modulować działanie kanałów wapniowych i potasowych, modyfikować rekrutację beta-arrestyny, wywoływać internalizację receptorów oraz pośrednio zmieniać przepływ substratów lipidowych, które zasilają również eikozanoidowe i inne zapalne szlaki. Model sieciowy lepiej niż wykres zajętości receptorów oddaje te konsekwencje międzyukładowe.
To także tutaj rozpada się dawne uproszczenie „CB1 w mózgu, CB2 w komórkach odpornościowych”. Rozmieszczenie jest stopniowalne, zależne od typu komórki i stanu fizjologicznego. CB1 występuje obficie w wielu populacjach neuronalnych, ale nie jednolicie w całym mózgu, a ponadto pojawia się w tkankach obwodowych istotnych dla bilansu energetycznego, nocycepcji i funkcji narządów. CB2 jest wzbogacony w liniach komórek układu odpornościowego, jednak nowsze badania nad OUN wyraźnie wykraczają poza pogląd, że jest on nieistotny dla biologii mózgu. Przegląd z 2026 roku opublikowany w Frontiers in Behavioral Neuroscience opisał to jako „aktualizację z ostatnich 3 lat”, podkreślając sygnalizację CB2 w kontekstach neurozapalnych i neurodegeneracyjnych. Nie chodzi o to, że CB2 nagle stał się „także receptorem mózgowym”. Chodzi o to, że znaczenie receptora zależy od tego, które komórki są aktywne, uszkodzone, objęte zapaleniem albo adaptujące się.
Analiza sieciowa pomaga wyjaśnić, dlaczego leki ukierunkowane na receptory często wywołują szersze efekty, niż oczekiwano. Phytocannabinoid aktywny wobec CB1, taki jak Delta-9-THC, endokannabinoid taki jak anandamid oraz syntetyczny agonista mogą wszystkie angażować ten sam receptor, a mimo to prowadzić do odmiennych efektów czasowych i przestrzennych. Liczy się klasa ligandu. Liczą się też skuteczność wewnętrzna, czas przebywania w miejscu wiązania, metabolizm i dostęp do różnych konformacji receptora.
CB1 i CB2 jako węzły o dużym znaczeniu
Integracyjne badanie sieciowe identyfikuje CB1 i CB2 jako węzły o dużym znaczeniu, nie dlatego, że działają samodzielnie, lecz dlatego, że znajdują się w punktach kontrolnych, gdzie krzyżuje się wiele szlaków. W terminologii sieciowej węzły o dużym znaczeniu to miejsca, w których lokalne zmiany mogą szeroko się propagować. W farmakologii są to receptory, których stan aktywacji może jednocześnie przekierowywać sygnalizację synaptyczną, odpowiedzi immunologiczne i programy komórkowego stresu.
CB1 jest wyraźniejszym przykładem tego, dlaczego „o dużym znaczeniu” nie znaczy „prosty”. Sygnalizuje głównie przez białka Gi/o, ale to dopiero początek. W zależności od ligandu i kontekstu CB1 może angażować beta-arrestyny, ulegać fosforylacyjnie zależnej desensytyzacji, internalizować się, recyklingować albo pozostawać trwale aktywny w błonie. Artykuł w American Journal of Psychiatry z 2025 roku argumentuje, że sygnalizacja wybiórcza przy CB1 jest prawdopodobną strategią terapeutyczną w schizofrenii, zaburzeniu, które według WHO dotyczy około 24 milionów ludzi na świecie. Taka propozycja ma sens tylko wtedy, gdy CB1 rozumie się jako węzeł sygnalizacyjny o rozdzielnych wyjściach, a nie jako przełącznik „włącz/wyłącz” dla odurzenia. Ligand faworyzujący jedną gałąź sygnalizacji nad inną może zachować część efektów terapeutycznych przy jednoczesnym ograniczeniu działań niepożądanych. Może, nie musi. Bias może zmniejszać ryzyko działań ubocznych, ale nie usuwa faktu, że CB1 znajduje się w obwodach kontrolujących poznanie, nagrodę, motorykę, przetwarzanie bólu i apetyt.
CB2 pokazuje podobny problem, ale z przeciwnej strony. Przez długi czas uznawano go za cel bezpieczniejszy, ponieważ słabiej wiązano go z ostrymi efektami psychoaktywnymi. To ujęcie jest zbyt optymistyczne. Ligandy ukierunkowane na CB2 mogą nadal dawać rozczarowujące lub mieszane wyniki, jeśli sygnalizacja istotna dla choroby zależy od stanu komórki, indukowalności receptora, napięcia zapalnego lub współdziałania z enzymami metabolicznymi. Najnowszy przegląd strukturalny w Frontiers in Chemical Biology, opublikowany w 2026 roku, podkreśla, że selektywność między „CB1 i CB2” zależy od różnic strukturalnych na poziomie receptora, które zmieniają wiązanie, skuteczność i regulację receptora. Badanie indeksowane w PubMed z 2025/2026 roku dotyczące selektywności podtypowej (PMID: 41962866) idzie dalej, argumentując, że selektywność jest dynamiczna i kształtowana przez zachowanie konformacyjne, a nie przez statyczne dopasowanie typu klucz-zamek. To mocny powód, aby przestać mówić o związkach selektywnych wobec CB1 lub CB2 tak, jakby selektywność była stałą cechą o stałych konsekwencjach biologicznych.
Powiązania między sygnalizacją receptorową a szlakami metabolicznymi
Najcenniejszym wkładem perspektywy sieciowej jest ponowne połączenie farmakologii receptorowej z metabolizmem lipidów. Sygnalizacja endokannabinoidowa jest zbudowana na chemii związanej z kwasem arachidonowym. Anandamid i 2-AG są wytwarzane i degradowane przez układy enzymatyczne, które wpływają również na dostępność bioaktywnych lipidów uczestniczących w zapaleniu i odpowiedzi tkankowej. Lek blokujący FAAH lub MAGL nie tylko „podnosi poziom endokannabinoidów”. On przemieszcza ruch sygnalizacyjny przez sieć metaboliczną.
To może być korzystne. Może też obrócić się przeciwko założonym efektom. Podwyższenie 2-AG może na przykład nasilać sygnalizację CB1 lub CB2 w niektórych przedziałach, a jednocześnie zmieniać szlaki związane z prostaglandynami w innych, poprzez odciąganie substratu lub dalsze utlenianie. Implikacja kliniczna jest prosta: strategie ukierunkowane na receptory i strategie ukierunkowane na enzymy mogą mieć skutki ogólnosystemowe, nawet jeśli ich projekt zakłada wąski cel. To jeden z powodów, dla których związki selektywne nie przełożyły się automatycznie na czyste profile kliniczne w bólu, chorobach psychiatrycznych czy zaburzeniach zapalnych, mimo dużej niezaspokojonej potrzeby terapeutycznej. Sam ból przewlekły dotyczy prawie 1 na 5 dorosłych w Stanach Zjednoczonych, według National Academies, a jednak farmakologia cannabinoid nie doprowadziła do powszechnie wiarygodnego rozwiązania receptorowego.
W tym ujęciu CB1 i CB2 lepiej rozumieć jako węzły sterujące osadzone w obwodach lipidowych, immunologicznych i synaptycznych. Ich podatność na działanie leków jest rzeczywista. Rzeczywista jest także ich zdolność do zaskakiwania.
CB1 jako cel lekowy: potencjał, ograniczenia i problem ośrodkowych działań niepożądanych
CB1 pozostaje jednym z najbardziej kuszących celów w neurofarmakologii, ponieważ znajduje się w strategicznym punkcie kontrolnym sygnalizacji synaptycznej. Prace Allyn Howlett z zakresu farmakologii receptorowej wykazały, że efekty wywoływane przez cannabinoid nie były niejasnym zjawiskiem błonowym, lecz biologią pośredniczoną przez receptor, a późniejsze odkrycie anandamidu przez Raphaela Mechoulama i Lumíra Hanuša nadało temu receptorowi endogenny system sygnalizacyjny. Ta historia ma znaczenie. Receptor, który pomaga regulować uwalnianie neuroprzekaźników w obwodach bólu, obwodach regulacji łaknienia, szlakach stresowych i sieciach nagrody, wydaje się wiarygodnym punktem uchwytu dla kilku trudnych zaburzeń jednocześnie. Jednocześnie stwarza pułapkę: gdy jeden receptor jest osadzony w tak wielu podstawowych funkcjach mózgu, szeroka modulacja rzadko pozostaje ograniczona do pożądanego efektu.
Problem ten nie ma charakteru akademickiego. World Health Organization szacuje, że w 2019 roku 200 milionów ludzi używało cannabis, czyli około 4% globalnej populacji w wieku 15–64 lat, więc farmakologia CB1 już teraz działa na skalę populacyjną poprzez kannabinoidy roślinne, zwłaszcza delta-9-THC. Mimo to zatwierdzonych leków z grupy cannabinoidów pozostaje niewiele: U.S. FDA podaje, że do 2025 roku zatwierdziła jeden produkt leczniczy pochodzący z cannabis oraz trzy produkty lecznicze związane z cannabis. Luka między powszechną ekspozycją a ograniczoną liczbą zatwierdzonych terapii mówi o CB1 coś istotnego. Biologia jest przekonująca. Farmakoterapia jest trudna.
Uzasadnienie terapeutyczne w bólu, apetycie i neuropsychiatrii
Atrakcyjność zaczyna się od miejsca działania CB1 i jego funkcji. CB1 jest silnie eksprymowany w wielu presynaptycznych zakończeniach w ośrodkowym układzie nerwowym, gdzie endocannabinoid są często wytwarzane na żądanie w komórkach postsynaptycznych i przemieszczają się wstecz przez synapsę, aby hamować uwalnianie neuroprzekaźników. Ten układ retrogradowy może osłabiać transmisję glutaminianergiczną, GABA-ergiczną i inne szlaki sygnałowe, zależnie od typu komórki i obwodu. W przypadku bólu jest to oczywiste uzasadnienie: skoro niemal 1 na 5 dorosłych w Stanach Zjednoczonych żyje z bólem przewlekłym, jak odnotowały National Academies w 2017 roku, receptor mogący zmniejszać transmisję nocyceptywną i modyfikować afektywny wymiar bólu z pewnością przyciąga uwagę.
CB1 nie jest jednak prostym przełącznikiem przeciwbólowym. W niektórych obwodach hamowanie uwalniania GABA może znosić hamowanie neuronów dalszego rzędu, zamiast je wyciszać. W innych dominują efekty hamowania glutaminergicznego. Zależy to od synapsy, stanu sieci i ligandu. Endocannabinoid, takie jak anandamid i 2-AG, są sygnałami krótkotrwałymi, lokalnie wytwarzanymi. Phytocannabinoid i syntetyczni agoniści wchodzą z odmienną kinetyką, odmienną skutecznością i często z dużo szerszym w czasie zaangażowaniem receptorowym. To rozróżnienie pomaga wyjaśnić, dlaczego stwierdzenie „aktywacja CB1 zmniejsza ból” jest zbyt uproszczone, by bezpiecznie prowadzić projektowanie kliniczne.
Drugim głównym uzasadnieniem jest apetyt. Sygnalizacja CB1 nasila przyjmowanie pokarmu i współdziała z obwodami podwzgórzowymi oraz mezolimbicznymi, które łączą homeostazę energetyczną z nagrodą. To czyniło antagonizm CB1 atrakcyjną strategią przeciwotyłościową, a agonizm CB1 lub pośrednie wzmocnienie tej sygnalizacji potencjalną drogą w kacheksji lub utracie apetytu. Logika ta nie była irracjonalna. Wynikała z podstaw fizjologii. Jednak apetyt jest nierozerwalnie związany z nastrojem, istotnością bodźców i przetwarzaniem stresu, a wszystkie te procesy są również kształtowane przez CB1.
Neuropsychiatria to obszar, w którym receptor staje się najbardziej interesujący i najbardziej niebezpieczny. CB1 moduluje obwody istotne dla dopaminy, hamowanie korowe, plastyczność hipokampa, uczenie strachu i reaktywność na stres. Daje mu to teoretyczne znaczenie w zaburzeniach lękowych, stanach pourazowych, depresji, zaburzeniach związanych z używaniem substancji psychoaktywnych i chorobach psychotycznych. American Journal of Psychiatry argumentował w 2025 roku, że biasowana sygnalizacja CB1 jest prawdopodobną strategią w schizofrenii, zaburzeniu dotykającym około 24 milionów ludzi na świecie według WHO. Znaczenie tej pracy nie polega na tym, że lek ukierunkowany na CB1 w schizofrenii jest gotowy. Polega na tym, że dziedzina odeszła od prostego agonizmu i antagonizmu w stronę farmakologii selektywnej względem szlaku, czerpiąc z szerszego repertuaru GPCR.
Dlaczego celowanie w CB1 jest atrakcyjne naukowo, ale trudne klinicznie
CB1 jest atrakcyjny naukowo dokładnie z tego samego powodu, dla którego jest trudny klinicznie: stanowi gęsty węzeł sygnalizacyjny. Na poziomie kanonicznym CB1 sprzęga się głównie z białkami Gi/o, zmniejszając aktywność cyklazy adenylanowej, modulując kanały jonowe i hamując uwalnianie neuroprzekaźników. Już to wystarczyłoby, by uczynić go ważnym celem. Ale receptor rekrutuje także beta-arrestiny, ulega desensytyzacji i internalizacji oraz może być kształtowany przez modulatory allosteryczne i ligandozależne stany konformacyjne. Strukturalny przegląd w Frontiers in Chemical Biology z 2026 roku jasno pokazuje to zarówno dla CB1, jak i CB2: selektywność i skuteczność ligandów wynikają ze структуральных różnic na poziomie receptora, które wpływają na wiązanie, wynik sygnalizacyjny i regulację receptora. Podobnie wyglądające kannabinoidy mogą więc wywoływać istotnie odmienną farmakologię.
To nie jest jedynie szczegół chemii medycznej. To bezpośrednio przekłada się na działania niepożądane. Ligand silnie promujący jedną aktywną konformację może powodować większą internalizację receptora, większą tolerancję lub inny profil behawioralny niż ligand stabilizujący inny stan. Badanie indeksowane w PubMed z lat 2025/2026 dotyczące selektywności względem podtypów argumentowało ponadto, że selektywność endocannabinoid jest dynamiczna, a nie stanowi stałego zdarzenia typu „klucz-zamek”, wzmacniając ideę, że zachowanie receptora zmienia się wraz z kontekstem konformacyjnym. Twórcy leków nie mają do czynienia z jednym celem CB1. Mają do czynienia z ruchomym zespołem stanów.
Dlatego właśnie bezpośredni agoniści i antagoniści CB1 tak często rozczarowywali. Silny agonizm ośrodkowy może upośledzać pamięć, uwagę, sprawność psychomotoryczną i regulację lęku, a jednocześnie angażować szlaki nagrody w sposób komplikujący ryzyko uzależnienia. Silny antagonizm ośrodkowy może rzeczywiście zmniejszać apetyt, ale może też zaburzać te same obwody buforowania stresu i regulacji afektu, które endogenne cannabinoid zwykle stabilizują. CB1 jest głęboko zaangażowany w funkcje poznawcze, nagrodę i regulację stresu. Każdy lek, który zbyt silnie pcha ten układ w jedną stronę, należy uznać za ryzykowny, dopóki nie zostanie udowodnione inaczej.
Biologia systemowa potwierdza tę ostrożność. Zintegrowana analiza sieciowa z lat 2025/2026 wskazała CB1 i CB2 jako wysoce wpływowe węzły w systemie endocannabinoid i połączyła sygnalizację receptorową z szerszymi szlakami metabolicznymi. Wynik ten przemawia przeciwko uproszczeniu opartego wyłącznie na receptorze. Zmiana CB1 nie przesuwa tylko jednej synapsy. Może propagować się przez sieci endokrynne, immunologiczne, metaboliczne i behawioralne.
Wnioski z podejść ośrodkowych i obwodowo ograniczonych
Najbardziej oczywistą lekcją z pierwszej ery rozwoju leków ukierunkowanych na CB1 jest to, że ekspozycja ośrodkowa często sprawia, iż skuteczność i ryzyko stają się nierozdzielne. Ośrodkowo aktywne agoniści CB1 mogą oddziaływać na obwody bólu i apetytu, ale ta sama penetracja do mózgu zwiększa prawdopodobieństwo efektów przypominających intoksykację, sedacji, zaburzeń poznawczych i działań niepożądanych psychiatrycznych. Ośrodkowo aktywne antagoniści CB1 pokazują odwrotny mechanizm niepowodzenia: mogą osiągać cele metaboliczne, a jednocześnie wywoływać nieakceptowalne efekty dotyczące nastroju i lęku, ponieważ blokują także endogenny ton cannabinoidowy w obwodach limbicznych.
Ta historia sprawiła, że ligandy CB1 o ograniczonym działaniu obwodowym stały się atrakcyjne. Idea jest prosta: jeśli część korzyści terapeutycznych zależy od CB1 w tkankach obwodowych, takich jak pierwotne włókna aferentne, jelito, wątroba, tkanka tłuszczowa lub inne miejsca poza OUN, to utrzymanie leku poza mózgiem może zachować użyteczne efekty przy jednoczesnym obniżeniu ośrodkowych działań niepożądanych. W bólu mogłoby to oznaczać wpływ na obwodowe mechanizmy nocyceptywne. W chorobie metabolicznej mogłoby to oznaczać ograniczenie lipogenezy lub modyfikację szlaków związanych z glukozą bez zaburzania nastroju czy poznania.
Logika jest słuszna, ale nie jest to rozwiązanie magiczne. Po pierwsze, obwodowa i ośrodkowa biologia CB1 nie są w stanach chorobowych czysto rozdzielne. Ból przewlekły, jedzenie i stres obejmują pętle łączące mózg i ciało. Po drugie, wykluczenie przez barierę krew-mózg jest problemem ilościowym, a nie binarnym. Nawet niewielka penetracja OUN może mieć znaczenie, zwłaszcza przy przewlekłym dawkowaniu. Po trzecie, nawet rzeczywiście obwodowo działający ligand CB1 może zawieść klinicznie, jeśli docelowa choroba silnie zależy od obwodów ośrodkowych lub jeśli szlaki kompensacyjne osłabiają skuteczność.
Nowsza strategia to nie tylko „pozostać poza mózgiem”. To „kształtować sygnał”. Obejmuje to częściowych agonistów, neutralnych antagonistów zamiast odwrotnych agonistów, modulatory allosteryczne oraz ligandy biasowane, preferujące wybrane szlaki downstream względem innych. Dyskusja w American Journal of Psychiatry z 2025 roku dotycząca biasowanej sygnalizacji CB1 w schizofrenii dobrze to ilustruje. Jeśli ligand może zachować wybrane sygnały terapeutyczne przy jednoczesnym ograniczeniu rekrutacji beta-arrestiny, desensytyzacji receptora lub niepożądanych efektów obwodowych, CB1 może stać się bardziej podatny na interwencję. Może. Nadal nie ma gwarancji, że bias szlaku obserwowany w systemach komórkowych będzie się dobrze przekładał na terapeutyczne okno u ludzi.
Obecne spojrzenie na CB1 jako cel lekowy nie jest więc ani lekceważące, ani romantyczne. CB1 jest atrakcyjny, ponieważ reguluje wzmocnienie synaptyczne dokładnie w tych obwodach, na które klinicyści chcą oddziaływać. Jest trudny, ponieważ te same obwody regulują pamięć, afekt, motywację i odporność na stres. Ligandy o ograniczonym działaniu obwodowym, strategie selektywnej sygnalizacji i lepsze zrozumienie strukturalne poprawiły logikę projektowania. Nie usunęły jednak podstawowego problemu. Receptor tak centralny dla funkcji mózgu rzadko toleruje farmakologię „tępą”.
CB2 jako cel leku: neurozapalenie, neurodegeneracja i modulacja odporności
CB2 stał się jednym z najbardziej kuszących celów w farmakologii cannabis z prostego powodu: wydaje się oferować dostęp do biologii immunologicznej i glejowej przy mniejszym ryzyku odurzenia, zaburzeń pamięci oraz potencjału nadużywania, które klasycznie wiążą się z silną aktywacją CB1. Ta atrakcyjność ma znaczenie wykraczające daleko poza akademickie mapowanie receptorów. WHO oszacowała, że w 2019 roku 200 milionów ludzi używało cannabis, czyli 4% światowej populacji w wieku 15–64 lata, a U.S. FDA nadal wymienia jedynie jeden produkt leczniczy pochodzący z cannabis i trzy produkty lecznicze związane z cannabis jako zatwierdzone w 2025 roku. Zainteresowanie jest duże. Tłumaczenie jest trudne.
Starszy skrót myślowy mówił, że CB1 to receptor mózgowy, a CB2 to receptor odpornościowy. To dziś zbyt uproszczone, by było użyteczne. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience, wprost ujęty jako „aktualizacja z ostatnich 3 lat”, traktuje CB2 jako istotny dla zaburzeń ośrodkowego układu nerwowego właśnie dlatego, że uczestniczy w mechanizmach neurozapalnych i neurodegeneracyjnych, w tym aktywacji mikrogleju, sygnalizacji cytokinowej oraz komunikacji neuron–glej. Kluczowa zmiana nie polega na tym, że CB2 nagle stał się obficie obecny wszędzie w mózgu. Tak się nie stało. Zmiana polega na tym, że niska podstawowa ekspresja w wielu obszarach CNS nie oznacza braku znaczenia funkcjonalnego, zwłaszcza gdy stany chorobowe mogą zmieniać ekspresję receptora, jego transport i sygnalizację.
Dlaczego CB2 jest często postrzegany jako bezpieczniejszy cel
Argument bezpieczeństwa dla CB2 zaczyna się od rozmieszczenia, ale na nim się nie kończy. CB2 jest wzbogacony w komórki odpornościowe i często ulega regulacji w górę w aktywowanym mikrogleju, naciekających makrofagach oraz innych populacjach komórek zapalnych. Dla porównania CB1 jest gęsto eksprymowany na zakończeniach presynaptycznych w korze, hipokampie, zwojach podstawy i móżdżku, gdzie silnie kształtuje uwalnianie neuroprzekaźników. Ta różnica anatomiczna ma znaczenie, ponieważ ligand, który głównie angażuje CB2, z mniejszym prawdopodobieństwem odtworzy klasyczny profil psychoaktywny związany z CB1.
„Mniejsze prawdopodobieństwo” nie znaczy jednak „niemożliwe”. Selektywność jest celem ruchomym. Przegląd z 2026 roku w Frontiers in Chemical Biology podkreśla, że selektywność „CB1 i CB2” zależy od różnic strukturalnych wpływających na wiązanie, skuteczność i regulację receptora, a niedawne badanie indeksowane w PubMed dotyczące selektywności podtypowej argumentuje, że rozróżnianie endocannabinoidów między podtypami receptorów odzwierciedla dynamikę konformacyjną, a nie stały model klucza i zamka. W praktyce oznacza to, że związek może wyglądać na selektywny wobec CB2 w jednym teście, a zachowywać się mniej czysto w innym, zwłaszcza gdy gęstość receptora, środowisko błonowe i dalsze transduktory są różne. Ten sam ligand może w jednym typie komórek wykazywać bias wobec sygnalizacji białka G, w innym rekrutować beta-arrestyny, a w warunkach zapalnych wykazywać odmienną kinetykę desensytyzacji lub internalizacji.
Ta złożoność mechanistyczna w rzeczywistości wzmacnia argument za ostrożnym rozwijaniem CB2. Jeśli celem terapeutycznym jest zmniejszenie wydzielania cytokin przez mikroglej, ograniczenie rekrutacji obwodowych komórek odpornościowych lub modyfikacja stresu metabolicznego gleju bez silnego zaburzania synaptycznego przekaźnictwa korowego, to CB2 pozostaje bardziej prawdopodobnym pierwszym receptorem do ukierunkowania. Bias w sygnalizacji CB1, jak argumentowano w artykule z 2025 roku w American Journal of Psychiatry dotyczącym schizofrenii, może ostatecznie poszerzyć okno terapeutyczne dla leków ukierunkowanych na CB1. Jest to jednak argument za mądrzejszą farmakologią CB1, a nie powód, by ignorować niższe ryzyko psychoaktywności podejść skoncentrowanych na CB2.
Bezpieczniejszy nie powinien jednak być mylony ze skuteczniejszym. Lek może nie wywoływać oczywistego odurzenia, a mimo to zawieść, ponieważ receptor jest w procesie chorobowym eksprymowany zbyt skąpo, zbyt zmiennie lub zbyt przejściowo, aby wywołać istotny efekt kliniczny. Zdarzało się to wielokrotnie w rozwoju leków przeciw neurozapoleniu, nie tylko w nauce o cannabis.
Dowody i ograniczenia w modelach chorób CNS
Przegląd w Frontiers in Behavioral Neuroscience przedstawia, dlaczego CB2 stale powraca w modelach choroby Alzheimera, choroby Parkinsona, stwardnienia rozsianego, urazowego uszkodzenia mózgu, udaru i bólu neuropatycznego: wszystkie te stany obejmują sygnalizację zapalną, reaktywność gleju, stres oksydacyjny i postępujące uszkodzenie tkanki. W wielu badaniach na gryzoniach agoniści CB2 zmniejszają mediatory prozapalne, takie jak TNF-α, IL-1β i IL-6, przesuwają mikroglej ku mniej szkodliwym fenotypom lub osłabiają infiltrację leukocytów. Czasami towarzyszy temu poprawa zachowania ruchowego, zmniejszenie rozmiaru ogniska uszkodzenia lub zachowanie markerów synaptycznych. To są realne sygnały, a nie artefakty do odrzucenia.
Ale obraz jest nierówny. Efekty często zależą od czasu. Agonista CB2 podany w czasie wczesnej fali zapalnej po urazie może działać zupełnie inaczej niż ten sam agonista podany w fazie przewlekłej degeneracji. Znaczenie ma także dawka oraz klasa ligandu. Endocannabinoidy, takie jak 2-AG i anandamid, nie są zamienne z fitocannabinoidami ani z syntetycznymi agonistami preferującymi CB2. Różnią się lokalne stężenia, rozkład metaboliczny i działania poza celem. Różnią się także preferencje szlaków sygnałowych.
Same modele chorobowe są kolejnym ograniczeniem. Transgeniczna mysz z chorobą Alzheimera, z patologią amyloidową i reaktywnym mikroglejem, nie jest chorobą Alzheimera u 74-letniego człowieka z mieszanym uszkodzeniem naczyniowym, zapalnym i proteostatycznym. Eksperymentalne autoimmunologiczne zapalenie mózgu i rdzenia jest użyteczne dla biologii stwardnienia rozsianego, ale odzwierciedla tylko część ludzkiego schorzenia. Pozytywne wyniki przedkliniczne ustanawiają więc wiarygodność, a nie dowód.
Istnieje też trwały problem pomiarowy w biologii CB2. Specyficzność przeciwciał od dawna stanowi słaby punkt, przez co twierdzenia o lokalizacji receptora są mniej wiarygodne, niż sugeruje wiele prac. Niska ekspresja w zdrowym mózgu, indukowalna ekspresja w chorobie oraz różnice gatunkowe dodatkowo komplikują interpretację. Część doniesień o „neuronalnym CB2” może być prawdziwa w określonych kontekstach, ale szerokie twierdzenia o powszechnej konstytutywnej ekspresji neuronalnego CB2 pozostają mniej pewne, niż kiedyś zakładano.
Tu sygnalizacja ma znaczenie równie duże jak anatomia. CB2 jest receptorowym GPCR sprzężonym z Gi/o, ale ten skrót ukrywa istotną zmienność: hamowanie cyklazy adenylanowej, modulacja szlaków MAPK, wpływ na kanały jonowe, rekrutacja beta-arrestyn, desensytyzacja receptora i internalizacja mogą wszystkie zmieniać netto wynik biologiczny. Ligand, który zmniejsza uwalnianie cytokin w hodowanych komórkach mikrogleju, może zawieść in vivo, jeśli szybko desensytyzuje receptory lub uruchamia niekorzystny program dominowany przez arrestynę. Niższe ryzyko odurzenia nie rozwiązuje tych problemów farmakologicznych.
Co sugeruje nowsza literatura dla przyszłych terapii
Nowsza literatura odchodzi od starego poszukiwania jednego „agonisty CB2 na zapalenie mózgu” i przesuwa się ku bardziej selektywnej, zależnej od kontekstu strategii. Przegląd strukturalny z 2026 roku oraz praca z 2025/2026 roku o selektywności podtypowej wspierają tę samą lekcję: ukierunkowanie na receptor będzie zależeć od stanu konformacyjnego, profilu skuteczności i kontekstu tkankowego, a nie tylko od nominalnego powinowactwa do podtypu. Projektowanie leków zmierza ku ligandom, które są selektywne funkcjonalnie, a nie wyłącznie pod względem wiązania.
Prawdopodobnie oznacza to trzy rzeczy. Po pierwsze, przyszłe leki CB2 będą wymagały lepszych „odcisków palca” sygnalizacji, w tym biasu białko G versus beta-arrestyn, czasu pozostawania receptora w kompleksie z ligandem oraz efektów zależnych od typu komórki w systemach ludzkich, a nie tylko w unieśmiertelnionych liniach testowych. Po drugie, stratyfikacja chorób będzie miała znaczenie. Terapia ukierunkowana na CB2 może działać najlepiej tam, gdzie amplifikacja zapalna jest głównym napędem patologii, a nie tylko jej produktem ubocznym. Po trzecie, myślenie o terapii skojarzonej jest nieuniknione. Integracyjna analiza sieci opublikowana w niedawnym badaniu indeksowanym w PubMed zidentyfikowała CB1 i CB2 jako wpływowe węzły w szerszych szlakach endocannabinoidowych i metabolicznych, co odpowiada rzeczywistości chorób CNS: zapalenie, stres mitochondrialny, sygnalizacja lipidowa i dysfunkcja synaptyczna są ze sobą splecione.
W określonych stanach zapalnych modulacja CB2 może zmniejszać szkodliwą sygnalizację immunologiczną, ograniczając jednocześnie znaczną część obciążenia psychoaktywnego związanego z CB1.Preliminary evidence
To nie czyni CB2 ślepą uliczką. Czyni go celem, który prawdopodobnie należy do farmakologii precyzyjnej, a nie do mitologii receptorów. Najsilniejszy obecny argument nie brzmi: „aktywacja CB2 leczy neurodegenerację”. Jest węższy: w określonych warunkach zapalnych, w określonych populacjach komórek i przy odpowiednich właściwościach ligandu modulacja CB2 może zmniejszać szkodliwą sygnalizację immunologiczną, jednocześnie unikając znacznej części psychoaktywnego obciążenia związanego z CB1. To wiarygodna hipoteza terapeutyczna. Nie jest jeszcze klinicznie zweryfikowaną zasadą.
Związki cannabis w CB1 i CB2: THC, CBD, minor cannabinoids i ligandy syntetyczne
cannabis nie działa przez jedną chemiczną substancję ani przez jeden mechanizm receptorowy. To stwierdzenie brzmi oczywiście, a jednak w dyskusji publicznej farmakologia nadal bywa sprowadzana do fałszywej pary: THC równa się CB1, CBD równa się CB2. Literatura nie potwierdza takiego skrótu myślowego. cannabis zawiera wiele phytocannabinoids, a także terpenes, flavonoids i inne składniki, podczas gdy organizm wytwarza własne endocannabinoids, takie jak anandamide i 2-arachidonoylglycerol, zidentyfikowane w pracach związanych z Raphaelem Mechoulamem, Lumírem Hanušem i innymi. Ponadto większość tego, co naukowcy wiedzą o aktywacji receptorów, skuteczności, selektywności, desensytyzacji i biasie sygnałowym, pochodzi z ligandów syntetycznych projektowanych jako sondy badawcze lub punkty wyjścia do opracowania leków, a nie wyłącznie ze związków roślinnych.
To rozróżnienie ma znaczenie, ponieważ CB1 i CB2 nie są biernymi zamkami czekającymi na klucz cannabinoid. Są to receptory sprzężone z białkiem G klasy A, o wielu stanach konformacyjnych, preferencjach sprzężenia i losach regulacyjnych. Przegląd strukturalny z 2026 roku w Frontiers in Chemical Biology jasno wskazuje, że selektywność ligandów wobec CB1 i CB2 zależy od różnic strukturalnych na poziomie receptora, które zmieniają wiązanie, skuteczność i regulację receptora, a nie tylko od tego, czy dana cząsteczka „trafiam” w jeden receptor czy drugi. Niedawne badanie indeksowane w PubMed dotyczące selektywności podtypów receptorów również dowodzi, że preferencja endocannabinoidów wobec CB1 versus CB2 wynika z dynamiki konformacyjnej, a nie z prostego dopasowania typu klucz-zamek. Gdy więc dwa cannabinoids wyglądają chemicznie podobnie, ale wywołują różne efekty w mózgu, tkance odpornościowej lub obwodach bólowych, nie jest to kwestia poboczna. To sedno farmakologii.
Biorąc pod uwagę, że World Health Organization oszacowała, iż w 2019 roku 200 milionów ludzi używało cannabis, czyli około 4% globalnej populacji w wieku 15–64 lata, takie rozróżnienia nie są akademicką ciekawostką. Kształtują one odurzenie, twierdzenia terapeutyczne, bezpieczeństwo oraz to, dlaczego jedne leki celujące w receptory odnoszą sukces, a inne zawodzą.
Jak THC oddziałuje z CB1 i przyczynia się do efektów psychoaktywnych
Delta-9-tetrahydrocannabinol, czyli THC, pozostaje centralnym związkiem do zrozumienia odurzenia wywoływanego przez cannabis, ponieważ bezpośrednio angażuje CB1 w ośrodkowym układzie nerwowym. To najczytelniejsze wyjaśnienie efektów psychoaktywnych cannabis na poziomie receptorowym, nawet jeśli „psychoaktywny” obejmuje kilka odrębnych zjawisk: zaburzenie percepcji czasu, wzmocnienie nagrody, zaburzenie pamięci, lękotwórczość u części użytkowników oraz upośledzenie koordynacji ruchowej. Prace farmakologiczne Allyn Howlett pomogły ustanowić to oparte na CB1 ujęcie i pozostaje ono fundamentalne.
Ale „THC aktywuje CB1” to dopiero początek. THC nie jest idealnym przełącznikiem typu on/off. Zwykle opisuje się je jako częściowy agonistę CB1, co oznacza, że jego skuteczność zależy od gęstości receptorów, kontekstu tkankowego i użytego testu sygnałowego. W regionach o wysokiej ekspresji CB1, takich jak kora, hipokamp, jądra podstawy i móżdżek, może to wystarczyć do wywołania silnych efektów ośrodkowych. Na poziomie synaptycznym CB1 jest często presynaptyczny, a jego aktywacja hamuje uwalnianie neuroprzekaźników. Oznacza to, że THC może osłabiać uwalnianie glutaminianu w jednym obwodzie, GABA w innym i wywoływać przeciwne skutki na poziomie sieciowym, zależnie od tego, które komórki eksprymują receptor i kiedy.
Dlatego położenia receptora nie można sprowadzać do hasła „CB1 w mózgu”. CB1 rzeczywiście występuje obficie w mózgu, ale nie równomiernie, nie statycznie i nie identycznie funkcjonalnie we wszystkich typach neuronów. W efekcie działanie THC jest zależne od obwodu neuronalnego. Euforia i wzmocnienie są częścią obrazu. Podobnie jak zaburzenia pamięci krótkotrwałej i sygnalizacja istotna dla psychozy u osób podatnych.
Ten ostatni punkt stał się ważniejszy, a nie mniej ważny. Artykuł z 2025 roku w American Journal of Psychiatry argumentował, że biased signaling CB1 jest możliwą strategią terapeutyczną w schizofrenii, właśnie dlatego, że biologii CB1 nie da się rozumieć jako jednowymiarowej aktywacji receptora. Schizofrenia dotyka około 24 milionów ludzi na świecie według WHO, a każda poważna dyskusja o THC na CB1 musi uwzględniać fakt, że ta sama rodzina receptorów, którą wiąże się z odurzeniem, jest również badana pod kątem agonizmu z biasem, rekrutacji beta-arrestin i projektowania leków selektywnych względem szlaków sygnałowych. THC może aktywować sygnalizację sprzężoną z Gi/o, ale CB1 ulega też fosforylacji, desensytyzacji i internalizacji, co ma konsekwencje dla tolerancji i odpowiedzi dalszych szlaków. Dawka i czas mają znaczenie. Powtarzana ekspozycja zmienia układ.
Syntetyczne ligandy CB1 pokazały to dawno temu. Związki takie jak CP55,940 i WIN55,212-2 są silnymi agonistami badawczymi, które w testach laboratoryjnych często wywołują silniejsze lub bardziej „czyste” odpowiedzi receptorowe niż THC. Rimonabant, inwersyjny agonista CB1, pokazał kierunek przeciwny: blokowanie CB1 lub obniżanie jego aktywności poniżej poziomu podstawowego mogło zmniejszać apetyt i masę ciała, ale działania niepożądane ze strony psychiki pogrzebały ten lek. Ta historia kliniczna jest przestrogą. Selektywne celowanie w CB1 może być farmakologicznie eleganckie, a mimo to medycznie rozczarowujące.
Dlaczego CBD nie pasuje do prostego modelu agonisty CB1/CB2
CBD jest cannabinoids, który najczęściej ulega spłaszczeniu do mitu. Rutynowo przedstawia się go jako „ten nieodurzający, działający przez CB2” albo jako bezpośredni przeciwbiegun THC na poziomie receptorowym. Żadne z tych twierdzeń nie jest prawdziwe.
| Klasa związku lub przykład | Jak artykuł opisuje zachowanie receptora | Kluczowy wniosek |
|---|---|---|
| THC | Częściowy agonista CB1 i CB2; centralny dla odurzenia poprzez CB1 | Oddziaływanie z CB1 pomaga wyjaśnić efekty psychoaktywne, ale wynik nadal zależy od kontekstu |
| CBD | Nie jest prostym agonistą CB1 ani CB2; może działać allosterycznie lub pośrednio | CBD nie jest prostym lustrzanym odbiciem THC w odniesieniu do CB2 |
| Kannabinoidy mniejszościowe | Zgłaszane efekty różnią się zależnie od testu i stężenia | Nie należy uogólniać zachowania receptora z jednego kannabinoidu na wszystkie |
| Ligandy syntetyczne | Często używane do określania skuteczności, selektywności i biasu | Związki sondujące mogą zachowywać się bardzo różnie od kannabinoidów roślinnych |
CBD nie zachowuje się jak prosty agonista CB1 lub CB2 w taki sposób, w jaki THC działa na CB1. Jego powinowactwo do miejsc ortosterycznych CB1 i CB2 jest względnie niskie, a duża część jego farmakologii wydaje się obejmować działania pośrednie, allosteryczne lub niezwiązane bezpośrednio z receptorami cannabinoid. W zależności od badanego układu CBD opisywano jako ujemny modulator allosteryczny CB1, co może zmieniać sposób sygnalizacji THC lub endocannabinoidów bez prostego włączania albo wyłączania receptora. To rozróżnienie jest kluczowe. Modulowanie kształtu receptora i wydajności sygnałowej nie jest tym samym co klasyczny agonizm.
CBD oddziałuje też z celami wykraczającymi poza CB1 i CB2, w tym z TRPV1, 5-HT1A, PPAR-gamma, szlakami związanymi z adenozyną oraz enzymami uczestniczącymi w utrzymaniu tonu endocannabinoid. Część efektów może odzwierciedlać zmienioną sygnalizację anandamide, a nie bezpośrednią stymulację receptora. To pomaga wyjaśnić, dlaczego profil kliniczny CBD nie daje się łatwo dopasować do starego skrótu receptorowego. U.S. FDA wskazuje, że do 2025 roku zatwierdziła jeden produkt leczniczy pochodzenia cannabis oraz trzy produkty lecznicze związane z cannabis; najlepiej ugruntowane zastosowanie medyczne CBD dotyczy padaczki, schorzenia dotykającego około 50 milionów ludzi na świecie, i tej drogi terapeutycznej nie da się wyjaśnić uproszczonym modelem „CBD wiąże CB2 i zmniejsza stan zapalny”.
Nawet w przypadku CB2 obraz jest szerszy niż dawny narracyjny schemat obwodowy. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience opisał „aktualizację z ostatnich 3 lat”, w której sygnalizacja CB2 zyskała uwagę w zaburzeniach ośrodkowego układu nerwowego dzięki powiązaniom z mechanizmami neurozapalnymi i neurodegeneracyjnymi. Nie oznacza to, że CBD jest po prostu lekiem CB2. Oznacza to, że sam receptor ma większy zasięg funkcjonalny, niż sugeruje to przekaz publiczny.
Znaczenie oddzielenia związków roślinnych od mitów receptorowych
Phytocannabinoids nie są wymienne i nie są też wymienne z ligandami syntetycznymi. Minor cannabinoids, takie jak cannabigerol (CBG), cannabinol (CBN), tetrahydrocannabivarin (THCV) i cannabichromene (CBC), wykazano jako oddziałujące z CB1 i CB2 w sposób zależny od testu i stężenia. Niektóre wykazują słaby agonizm, inne częściowy agonizm, jeszcze inne antagonizm w określonych warunkach, a część wykazuje istotną aktywność wobec celów niezwiązanych z cannabinoid. THCV jest dobrym przykładem: w niektórych układach opisywano je jako antagonistę CB1 lub neutralnego antagonistę w niskich dawkach, a w innych jako częściowego agonistę przy wyższych dawkach. To nie jest niespójność dla samej niespójności. To odzwierciedlenie farmakologii receptora zależnej od kontekstu.
Narzędzia badawcze wyostrzają ten punkt. endocannabinoids, phytocannabinoids i ligandy syntetyczne mogą aktywować „CB1 i CB2”, ale nie stabilizują tych samych konformacji receptora ani nie wywołują tego samego biasu względem białek G i beta-arrestin. Przegląd strukturalny z 2026 roku oraz najnowsze analizy sieciowe indeksowane w PubMed kierują pole badań z dala od mitologii receptorowej ku farmakologii systemowej. Receptory funkcjonują w obrębie swoistych dla komórki sieci sygnałowych, szlaków metabolicznych i mechanizmów transportu wewnątrzkomórkowego. Profil kliniczny ligandu wynika z całego tego środowiska, a nie z etykiety typu „CB2 przeciwzapalny” albo „CBD równoważy THC”.
Dla wiki o cannabis to właśnie ta granica ma największe znaczenie: THC ma podstawowe znaczenie dla odurzenia mediowanego przez CB1, ale farmakologia cannabis na tym się nie kończy, a CBD nie jest prostym lustrzanym odpowiednikiem THC wobec CB2. Związki roślinne rozpoczynają opowieść. Stan receptora, dystrybucja tkankowa, skuteczność, bias i regulacja decydują o tym, jak się ona kończy.
Schizofrenia, ryzyko psychozy i to, co biologia CB1 implikuje, a czego nie implikuje
Dlaczego schizofrenia pojawia się w rozmowie o receptorze
Schizofrenia ma tu znaczenie, ponieważ CB1 nie jest po prostu kolejnym receptorem na długiej liście celów. Jest jednym z najliczniejszych receptorów sprzężonych z białkiem G w mózgu, umiejscowionym na zakończeniach presynaptycznych, gdzie może osłabiać uwalnianie neuroprzekaźników i przebudowywać aktywność sieciową w obwodach już wiązanych z psychozą: korze, hipokampie, prążkowiu, ciele migdałowatym. Gdy receptor znajduje się tak blisko kontroli obwodów glutaminergicznych, GABA-ergicznych, dopaminergicznych i odpowiedzi na stres, badacze psychiatryczni zwracają na niego uwagę.
Skala zdrowia publicznego sprawia, że jest to coś więcej niż pytanie akademickie. World Health Organization szacuje, że cannabis było używane przez 200 milionów osób w 2019 roku, czyli przez 4% globalnej populacji w wieku 15–64 lata. WHO szacuje również, że schizofrenia dotyka około 24 milionów osób na świecie, czyli mniej więcej 1 na 300 osób. To nie są małe, odizolowane populacje. Nakładają się one w rzeczywistych poradniach, na oddziałach ratunkowych i w długoterminowych badaniach kohortowych. Każda biologia receptora, która może rzucić światło na ryzyko psychozy, rozwój leków przeciwpsychotycznych albo jedno i drugie, zasługuje na poważną analizę.
To jest translacyjne tło dla artykułu z 2025 roku w American Journal of Psychiatry, argumentującego, że stronnicze sygnalizowanie CB1 jest prawdopodobną strategią terapeutyczną w schizofrenii. Główna idea jest łatwa do sformułowania i łatwa do nadużycia. Łatwa do sformułowania: CB1 nie zachowuje się jak prosty przełącznik włącz/wyłącz, a różne ligandy mogą stabilizować różne konformacje receptora, przesuwając sygnalizację w kierunku określonych szlaków wewnątrzkomórkowych lub od nich odchodząc. Łatwa do nadużycia: jeśli CB1 jest istotny dla badań nad leczeniem schizofrenii, niektórzy czytelnicy wysuwają fałszywy wniosek, że „aktywacja cannabinoid pomaga w schizofrenii” albo że używanie cannabis ma więc z definicji charakter leczniczy. Artykuł nie wspiera takiego skoku logicznego.
Historycznie nauka o cannabinoid utknęła w skrótach myślowych. CB1 przedstawiano jako „receptor mózgowy”, CB2 jako „receptor immunologiczny”, a efekty psychoaktywne przypisywano niemal wyłącznie zajęciu CB1. To ujęcie było zawsze niepełne, a do 2025 roku jest już po prostu zbyt uproszczone. Kluczowe prace Allyn Howlett ustanowiły farmakologię receptorów cannabinoid, a Mechoulam i Hanus pomogli zdefiniować biologię endocannabinoid poprzez anandamid. Od tego czasu farmakologia receptorów przesunęła się od samej obecności receptora do stanu receptora. To ma znaczenie dla schizofrenii, ponieważ patofizjologia nie jest powodowana samą ekspresją receptora. Powstaje z czasu, typu komórki, kontekstu obwodowego i odpowiedzi wewnątrzkomórkowej.
Nowsza literatura wzmacnia ten punkt. Przegląd z 2026 roku w Frontiers in Chemical Biology wyjaśnia, że różnice strukturalne między CB1 i CB2 kształtują wiązanie liganda, skuteczność, sygnalizację i regulację receptora. Badanie indeksowane w PubMed z 2025/2026 roku dotyczące selektywności podtypów argumentuje, że selektywność endocannabinoid może wynikać z dynamiki konformacyjnej, a nie ze statycznego modelu klucz–zamek. Inna praca z zakresu analizy sieciowej z 2025/2026 roku identyfikuje CB1 i CB2 jako wysoce wpływowe węzły w całym systemie endocannabinoid, powiązane ze szlakami metabolicznymi i sygnałowymi, a nie z izolowanymi „pudełkami receptorowymi”. Dla badań nad schizofrenią oznacza to, że należy przestać pytać wyłącznie „czy związek trafia w CB1?” i zacząć pytać „które stany CB1, w jakich komórkach, przy jakich warunkach ekspozycji?”
Stąd bierze się kliniczna powaga. Nie z haseł o receptorach. Z precyzji mechanistycznej.
Stronnicza sygnalizacja versus bezpośrednia aktywacja receptora
Bezpośrednia aktywacja CB1 jest słabym modelem rozumowania terapeutycznego. Spłaszcza zbyt wiele zmiennych do jednego słowa: aktywacja. Endogenne ligandy, takie jak anandamid i 2-AG, fitocannabinoids, takie jak delta-9-THC, oraz syntetyczne agoniści nie zachowują się identycznie wobec CB1, nawet jeśli wszystkie są ligandami. Mogą różnić się powinowactwem, skutecznością, czasem przebywania przy receptorze, ekspozycją regionalną i preferencją szlaku sygnałowego. Mogą też różnić się siłą rekrutacji białek Gi/o względem beta-arrestin, szybkością wywoływania desensytyzacji oraz stopniem internalizacji receptora po powtarzanym dawkowaniu.
To rozróżnienie jest kluczowe dla argumentu AJP dotyczącego schizofrenii. Ligand, który delikatnie kieruje CB1 w stronę jednego programu wewnątrzkomórkowego, nie musi przypominać liganda, który wywołuje szeroką aktywację receptora w wielu obszarach mózgu. W farmakologii GPCR taka różnica może oddzielać lek użyteczny od leku nietolerowanego. CB1 jest szczególnie wrażliwy na ten problem, ponieważ ten sam receptor może ograniczać transmisję pobudzającą w jednej synapsie, hamować transmisję hamującą w innej i zmieniać plastyczność downstream w sposób niebędący funkcjonalnie równoważnym. Niewielkie różnice molekularne mogą dawać duże konsekwencje na poziomie całych układów.
THC jest oczywistym przykładem tego, dlaczego „cel CB1” nie oznacza „terapia CB1”. THC jest fitocannabinoid z właściwościami częściowego agonisty CB1, ale odczuwalne i kliniczne efekty THC nie są czystym odczytem jednego ramienia sygnalizacji. Znaczenie ma dawka. Znaczenie ma wiek ekspozycji. Znaczenie ma powtarzana ekspozycja. Znaczenie ma także stosunek THC do innych składników oraz to, czy dana osoba ma podatność psychiatryczną. Ostre efekty podobne do intoksykacji, lęk, zmiana percepcji i przejściowe objawy psychotyczne u części użytkowników nie oznaczają, że wszystkie terapie ukierunkowane na CB1 są skazane na porażkę. Oznaczają natomiast, że bezrefleksyjna stymulacja CB1 nie jest skrótem do leczenia przeciwpsychotycznego.
Stronniczy agonizm jest atrakcyjny właśnie dlatego, że próbuje uniknąć tej bezpośredniości. Nadzieja terapeutyczna nie brzmi „więcej CB1”. Brzmi „właściwy profil sygnalizacji CB1”. Może to oznaczać preferowanie szlaków związanych ze stabilizacją obwodów, przy jednoczesnym ograniczaniu szlaków powiązanych z niepożądanymi efektami poznawczymi lub psychotomimetycznymi, tolerancją albo downregulation receptora. Czy da się to osiągnąć u pacjentów, pozostaje otwartym pytaniem. Koncepcja jest wiarygodna. Nie jest dowiedzionym sukcesem klinicznym.
CB2 również należy uwzględnić w tym obrazie, choć nie jako uproszczone rozwiązanie. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience opisuje „aktualizację z ostatnich 3 lat”, w której sygnalizacja CB2 zyskała uwagę w zaburzeniach ośrodkowego układu nerwowego z powodu powiązań neurozapalnych i neurodegeneracyjnych. Osłabia to dawny pogląd, że CB2 jest nieistotny dla chorób mózgu. Nie przywraca jednak starego binarnego podziału przez jego odwrócenie. Schizofrenia nie jest wyłącznie stanem zapalnym, a CB2 nie jest po prostu przyciskiem przeciwzapalnym. Każdy poważny model translacyjny musi uwzględniać jednocześnie interakcje neuron–glej, sygnalizację immunologiczną i dysfunkcję obwodową.
Czego nie należy wnioskować
- Nie jest to dowód korzyści To, że CB1 jest celem leku, nie oznacza, że używanie konopi chroni przed schizofrenią lub psychozą.
- Nie obala ryzyka Mechanistyczne badania receptorów nie znoszą obaw epidemiologicznych dotyczących ekspozycji na wysokie THC u osób podatnych.
- Nie jest to zamienność Endokannabinoidów, fitokannabinoidów i ligandów syntetycznych nie należy traktować jako tego samego rodzaju narażenia.
- Nie jest to ogólna aprobata Istniejące zatwierdzenia FDA nie rozstrzygają kwestii bezpieczeństwa psychiatrycznego ani skuteczności szerokiego stosowania kannabinoidów.
Czego czytelnicy nie powinni wywnioskować z badań mechanistycznych
Po pierwsze, czytelnicy nie powinni wyciągać wniosku, że skoro CB1 jest celem terapeutycznym, to używanie cannabis jest więc ochronne wobec schizofrenii lub psychozy. Logika celu lekowego nie działa w ten sposób. Receptory beta są celami leków; to nie znaczy, że każdy stymulant jest terapią. Receptory opioidowe są celami leków; to nie znaczy, że każdy agonizm opioidowy jest korzystny. Ta sama zasada obowiązuje tutaj.
Po drugie, badania mechanistyczne nie znoszą obaw epidemiologicznych. Badania nad stronniczością receptora i projektowaniem liganda współistnieją z dowodami, a nie przeciwko nim, że ekspozycja na cannabis może zwiększać ryzyko psychozy u części osób, zwłaszcza przy produktach o wysokiej zawartości THC, wczesnej inicjacji lub ciężkim używaniu. Mechanizm receptorowy może wyjaśniać, dlaczego niektóre związki można projektować tak, by dawały lepsze wyniki. Nie uzasadnia on szerokich twierdzeń publicznych, że „aktywacja CB1 jest dobra dla schizofrenii”.
Po trzecie, czytelnicy nie powinni traktować endocannabinoid, fitocannabinoids i ligandów syntetycznych jako wymiennych. Anandamid nie jest THC. Obwodowo ograniczony modulator CB1 nie jest palonym cannabis. Niskoefektywny, stronniczy względem szlaku związek badawczy nie jest konsumenckim produktem o wysokiej ekspozycji. Te kategorie mają znaczenie, ponieważ zachowanie receptora zależy od klasy liganda, dawki, drogi podania, czasu i ekspozycji tkankowej.
Na koniec, istnienie zatwierdzonych leków związanych z cannabis nie rozstrzyga kwestii psychiatrycznych. Według stanu na 2025 rok FDA podaje, że zatwierdzono jeden lek pochodzący z cannabis i trzy leki związane z cannabis. Ten fakt pokazuje, że farmakologia cannabinoid może stać się medycyną w ściśle określonych warunkach. Nie oznacza jednak, że nieprzetworzona stymulacja receptora jest bezpieczna, skuteczna ani odpowiednia w schizofrenii.
Właściwy wniosek jest bardziej rygorystyczny i mniej pocieszający. Biologia CB1 ma znaczenie dla psychozy, ponieważ receptor jest głęboko osadzony w obwodach mózgowych kształtujących istotność bodźców, poznanie i hamowanie. To znaczenie wspiera ostrożny projekt leków, a nie swobodne ekstrapolacje. W badaniach nad schizofrenią farmakologia receptorów jest jednocześnie mapą możliwości i obciążeń.
Co nauka o receptorach oznacza dla przyszłych leków cannabinoid
Medycyna cannabinoid odchodzi od starej idei, że użyteczny lek musi po prostu „trafić w CB1” albo „trafić w CB2”. Ten skrót myślowy był zawsze niepełny. Obecnie jest aktywnie mylący. CB1 i CB2 nie są prostymi przełącznikami włącz/wyłącz podzielonymi jasno na układ nerwowy i układ immunologiczny. Są receptorami o nakładających się, ale nierównomiernych rozkładach między typami komórek, dynamicznych konformacjach, odmiennych preferencjach sprzężenia i zmieniających się rolach w stanach chorobowych. Ma to znaczenie, ponieważ ten sam ligand może wyglądać jak terapeutyczny w jednej tkance, jak odurzający w innej, a w badaniu klinicznym może być nieskuteczny, jeśli populacja chorobowa jest zbyt szeroka albo schemat dawkowania prowadzi do desensytyzacji.
Stawka zdrowia publicznego nie jest błaha. Światowa Organizacja Zdrowia oszacowała, że w 2019 roku 200 milionów ludzi używało cannabis, co odpowiadało 4% globalnej populacji w wieku 15–64 lata. Mimo to amerykańska FDA nadal odnotowuje, że do 2025 roku zatwierdzono tylko jeden produkt leczniczy pochodzący z cannabis oraz trzy produkty lecznicze związane z cannabis. Ta luka między powszechną ekspozycją a ograniczoną liczbą zatwierdzonych terapii jest jednym z powodów, dla których farmakologia receptorowa ma tak duże znaczenie. Innym jest obciążenie chorobami: padaczka dotyczy około 50 milionów osób na świecie, schizofrenia około 24 milionów, a przewlekły ból dotyka niemal 1 na 5 dorosłych w Stanach Zjednoczonych. Jeśli biologia cannabinoidów ma doprowadzić do powstania lepszych leków, nie stanie się to poprzez prymitywną aktywację receptorów.
Fundamentalne prace Raphaela Mechoulama, Lumíra Hanuša i Allyn Howlett stworzyły nowoczesną dziedzinę, identyfikując endocannabinoid signaling oraz receptor pharmacology. Kolejny etap wygląda inaczej. Chodzi już mniej o dowodzenie, że CB1 i CB2 istnieją, a bardziej o naukę precyzyjnego sterowania nimi.
| Koncepcja projektowa | Co ma poprawić | Dlaczego według artykułu ma to znaczenie |
|---|---|---|
| Selektywność podtypowa | Bardziej precyzyjnie preferować CB1 lub CB2 | Sam podtyp nie gwarantuje skuteczności ani bezpieczeństwa |
| Dostrajanie skuteczności | Unikać zbyt silnej aktywacji receptora | Różne stany aktywne mogą zmieniać korzyści, tolerancję i działania niepożądane |
| Sygnalizacja biegunowana | Faworyzować jeden szlak wewnątrzkomórkowy nad innym | Może pomóc oddzielić korzystne efekty od ograniczeń |
| Modulacja allosteryczna | Dostroić odpowiedź receptora na endogenne ligandy | Mogłoby to zachować bardziej prawidłową sygnalizację przestrzenną i czasową |
| Ograniczenie tkankowe | Ograniczyć ekspozycję do wybranych narządów lub uniknąć przenikania do mózgu | Przydatne, gdy dominują działania niepożądane ośrodkowe |
Selektywność, skuteczność, modulacja allosteryczna i bias
Najważniejsza lekcja z najnowszej nauki o receptorach brzmi: „wiązanie” to nie to samo co „efekt”. Ligand może preferować CB1 względem CB2 albo odwrotnie, ale nawet to jest dopiero początek. Może też różnić się skutecznością, czyli tym, jak silnie stabilizuje aktywne stany receptora. Może faworyzować sygnalizację Gi/o zamiast rekrutacji beta-arrestin, albo bardziej promować internalizację receptora niż utrzymującą się sygnalizację z powierzchni komórki. Takie różnice mogą dramatycznie zmieniać wyniki.
Przegląd z 2026 roku opublikowany w Frontiers in Chemical Biology dotyczący struktury receptora cannabinoidowego jasno pokazuje tę kwestię: selektywność podtypowa między CB1 a CB2 zależy od różnic strukturalnych na poziomie receptora, które kształtują wiązanie liganda, skuteczność sygnalizacji i regulację receptora. Dlatego chemicznie podobne cannabinoids nie zachowują się identycznie. Fitocannabinoid, endocannabinoid, taki jak anandamid, oraz ligand syntetyczny mogą wszystkie oddziaływać z tą samą rodziną receptorów, a jednocześnie wywoływać różne profile efektów downstream. Projektowanie leków, które ignoruje skuteczność i bias sygnalizacyjny, jest dziś już niezgodne z dowodami.
CB1 jest najjaśniejszym przykładem. Często opisuje się go jako receptor odpowiedzialny za efekty psychoaktywne, co jest prawdą, ale niepełną. CB1 reguluje także transmisję synaptyczną, przetwarzanie bólu, apetyt, obwody stresowe i sygnalizację korową. Artykuł w American Journal of Psychiatry z 2025 roku argumentuje, że biased CB1 signaling jest prawdopodobną strategią terapeutyczną w schizofrenii. To uderzająca zmiana w tej dziedzinie. Schizofrenia dotyka około 24 milionów ludzi na świecie, a artykuł nie traktuje CB1 jako receptora, którego należy za wszelką cenę unikać. Zamiast tego ujmuje CB1 jako cel, którego patologiczne i terapeutyczne konsekwencje mogą zależeć od tego, które szlaki wewnątrzkomórkowe preferuje dany ligand. To klasyczna farmakologia GPCR, zastosowana teraz poważnie do terapii opartych na cannabinoidach.
CB2 przeszedł podobną rewaluację. Przegląd z 2026 roku w Frontiers in Behavioral Neuroscience opisuje „aktualizację z ostatnich 3 lat”, wskazując rosnące zainteresowanie sygnalizacją CB2 w zaburzeniach ośrodkowego układu nerwowego, zwłaszcza tam, gdzie biorą udział mechanizmy neurozapalne i neurodegeneracyjne. Nie oznacza to, że CB2 jest magicznym celem przeciwzapalnym. Oznacza to, że dawne twierdzenie, iż CB2 jest nieistotny dla mózgu, nie pasuje już do literatury. Ekspresja może być niska w warunkach wyjściowych, ale wzrastać w określonych populacjach komórek lub w kontekstach chorobowych. Lek ukierunkowany na CB2 może więc w dużym stopniu zależeć od czasu, patologii i stanu tkanki.
Modulacja allosteryczna może stać się jednym z najbardziej użytecznych sposobów wykorzystania tych różnic. Agoniści ortosteryczni często wywołują szeroką aktywację receptora, co zwiększa ryzyko działań niepożądanych, tolerancji i słabej selektywności kontekstowej. Z kolei modulatory allosteryczne mogą regulować sposób odpowiedzi receptorów na endogenne ligandy zamiast wymuszać maksymalną aktywację. W teorii mogłoby to zachować przestrzenny i czasowy charakter normalnego endocannabinoid signaling. To atrakcyjna strategia dla CB1, gdzie bezpośrednia agonistyka często napotyka centralne działania niepożądane, oraz dla CB2, gdzie chorobowo związana nadekspresja może sprawiać, że modulacja zależna od kontekstu jest bardziej pożądana niż stała stymulacja. Ale „atrakcyjne” nie znaczy „udowodnione”. Wiele eleganckich idei allosterycznych in vitro zawodzi, gdy pojawiają się rezerwa receptorowa, heterogenność tkankowa i farmakokinetyka.
Dlaczego struktura i biologia sieciowa kierują dziś projektowaniem leków
Ciężar gatunkowy tej dziedziny przesunął się od prostego mapowania receptorów do dynamicznej analizy stanów receptorowych. Tu właśnie znaczenie ma biologia strukturalna. Przegląd z 2026 roku w Frontiers oraz niedawne badanie indeksowane w PubMed dotyczące dynamicznego mechanizmu selektywności podtypowej odchodzą od modelu klucza i zamka. Selektywność nie jest już tylko kwestią tego, że jeden ligand lepiej pasuje do jednej kieszeni receptora. Receptory oddychają. Ligandy stabilizują całe zespoły konformacji. Endocannabinoids mogą osiągać preferencję podtypową dzięki dynamicznym interakcjom, które zmieniają sposób przechodzenia CB1 lub CB2 między stanami nieaktywnym, aktywnym i kompetentnym do sygnalizacji.
Ta zmiana myślenia ma praktyczne konsekwencje. Chemik medyczny nie pyta już tylko: „Czy ten związek wiąże CB1 lub CB2?” Lepsze pytania brzmią: Jakie konformacje stabilizuje? Czy promuje Gi/o bardziej niż beta-arrestin? Czy wywołuje szybką internalizację? Czy zachowuje się inaczej w neuronach, mikrogleju, hepatocytach lub obwodowych nocyceptorach? Czy można go ograniczyć do obwodu? Czy współdziała z endogennym tonusem ligandowym, który wzrasta tylko podczas zapalenia lub uszkodzenia?
Biologia sieciowa rozszerza to jeszcze dalej. Niedawna integracyjna analiza sieciowa indeksowana w PubMed zidentyfikowała CB1 i CB2 jako wysoce wpływowe węzły w endocannabinoid system i powiązała sygnalizację receptorową z szerszymi szlakami metabolicznymi. Ma to znaczenie, ponieważ farmakologia endocannabinoidów nigdy nie dotyczy wyłącznie receptorów. Synteza, transport i degradacja anandamidu oraz 2-AG kształtują ekspozycję receptorów. Podobnie działają metabolizm lipidów, mediatory zapalne oraz wzajemne oddziaływania z innymi systemami GPCR i kanałów jonowych. Ligand, który wygląda czysto w teście na rekombinowanym układzie, może zachowywać się zupełnie inaczej w żywej tkance, gdzie FAAH, MAGL, szlaki prostaglandyn i lokalna gęstość receptorów są zmienne.
To ujęcie systemowe pomaga też wyjaśnić, dlaczego translacja kliniczna była nierówna. Niektóre porażki prawdopodobnie wynikały ze złych związków. Inne z nadmiernie uproszczonych modeli chorób. Jeśli badanie rekrutuje pacjentów, których objawy mają wspólną nazwę, ale nie wspólny sygnatur endocannabinoid, średni wynik może wyglądać rozczarowująco, nawet jeśli biologicznie zdefiniowana podgrupa mogłaby odnieść korzyść. Przyszły sukces będzie zależał nie tylko od chemii, ale także od stratyfikacji: którzy pacjenci, jaki etap choroby, jakie tkanki, jaki stan receptora, jaki endogenny tonus.
Realistyczne oczekiwania wobec kolejnej generacji terapii
Kolejna generacja leków opartych na cannabinoidach prawdopodobnie będzie bardziej wąska, bardziej selektywna i mniej romantyczna, niż sugeruje debata publiczna. To dobra wiadomość. Lepsze leki często powstają wtedy, gdy dziedzina rezygnuje z wyjaśnień typu „jeden rozmiar dla wszystkich”.
Już teraz widać pewne postępy. Ograniczenie CB1 do obwodu pozostaje atrakcyjne w bólu lub wskazaniach metabolicznych, ponieważ ma na celu zmniejszenie centralnych działań niepożądanych. Związki ukierunkowane na CB2 pozostają prawdopodobne w stanach zapalnych, neuropatycznych i neuroimmunologicznych, zwłaszcza jeśli ekspresja związana z chorobą tworzy okna terapeutyczne. Ligandy CB1 o biased signaling mogą okazać się przydatne w chorobach neuropsychiatrycznych, jeśli potrafią oddzielić pożądane efekty sieciowe od niepożądanej intoksykacji, lęku lub tolerancji. Modulatory allosteryczne mogą oferować subtelniejszą kontrolę niż bezpośredni agoniści. Podejścia ukierunkowane na enzymy, które pośrednio zmieniają ton endocannabinoidów, nadal mogą być wartościowe, choć niosą własne ryzyka i nie omijają złożoności receptorowej.
Mimo to realizm jest niezbędny. Selektywność nie gwarantuje sukcesu. Wysoce selektywny agonista CB2 może nadal zawieść, jeśli ekspresja CB2 w docelowej chorobie jest zbyt zmienna albo jeśli sygnalizacja downstream nie jest rzeczywistym czynnikiem napędzającym objawy. Bias mierzony w jednym teście może nie przewidywać biasu w ludzkiej tkance. Obrazy strukturalne są potężne, ale pozostają obrazami migawkowymi. Ograniczenie do określonej tkanki może zmniejszyć jedne ograniczenia, a stworzyć inne. I fitocannabinoidów nie należy traktować jako substytutów całej farmakologii cannabinoidów; endocannabinoids, związki roślinne i ligandy syntetyczne mogą wywoływać bardzo różne zachowania receptorowe zależnie od dawki, czasu i kontekstu sygnalizacyjnego.
Zatem trzeźwa prognoza brzmi tak: przyszła medycyna oparta na cannabinoidach prawdopodobnie poprawi się dzięki większej precyzji, a nie dzięki silniejszej, szerokiej aktywacji. Najbardziej wiarygodne kierunki łączą selektywność receptorową, bias sygnalizacyjny, kontrolę allosteryczną, ukierunkowanie tkankowe oraz lepszą stratyfikację pacjentów. To, co już wiadomo, jest znaczące: CB1 i CB2 są dynamicznymi węzłami sygnalizacyjnymi, stan strukturalny ma znaczenie i kontekst chorobowy ma znaczenie. To, co pozostaje niepewne, to czy ta mechanistyczna jasność przełoży się na powtarzalne korzyści kliniczne w bólu, chorobach psychiatrycznych, padaczce, neurodegeneracji i chorobach zapalnych. Dziedzina nadal ma duże znaczenie medyczne, ponieważ niezaspokojona potrzeba jest ogromna, biologia jest realna, a dawny uproszczony model wreszcie ustąpił miejsca farmakologii zdolnej wyjaśnić zarówno obietnicę, jak i rozczarowanie.
Źródła
- [1]Cannabis (marijuana). WHO Fact Sheet, 2024. https://www.who.int/news-room/fact-sheets/detail/cannabis-(marijuana)
- [2]Epilepsy. WHO Fact Sheet, 2024. https://www.who.int/news-room/fact-sheets/detail/epilepsy
- [3]Schizophrenia. WHO Fact Sheet, 2022. https://www.who.int/news-room/fact-sheets/detail/schizophrenia
- [4]FDA Regulation of Cannabis and Cannabis-Derived Products, Including Cannabidiol (CBD). FDA Public Health Focus, 2025. https://www.fda.gov/news-events/public-health-focus/fda-regulation-cannabis-and-cannabis-derived-products-including-cannabidiol-cbd
- [5]The Health Effects of Cannabis and Cannabinoids: The Current State of Evidence and Recommendations for Research. National Academies Press, 2017. https://nap.nationalacademies.org/catalog/24625/the-health-effects-of-cannabis-and-cannabinoids-the-current-state