






Porque CB1 e CB2 são importantes na ciência do cannabis
GPCR Recetor acoplado à proteína G: um recetor de membrana que muda de forma após a ligação do ligando e sinaliza através de parceiros intracelulares, como proteínas G e beta-arrestinas.
O que determina a resposta do recetor
- Localização O tecido, o tipo celular e a posição subcelular moldam a resposta.
- Identidade do ligando Endocanabinoides, fitocanabinoides e ligandos sintéticos não desencadeiam estados de sinalização idênticos.
- Parceiros disponíveis Células diferentes oferecem proteínas G, cinases e beta-arrestinas distintas.
- Padrão de exposição A duração do sinal e a estimulação repetida afetam a dessensibilização e a internalização.
- Polarização da via Um ligando pode favorecer a sinalização por proteína G, o recrutamento de beta-arrestina ou outros resultados.
A linguagem pública tem, há muito, reduzido a biologia dos cannabinoid a uma divisão simplista: o CB1 explica o “high”, o CB2 trata da inflamação algures fora do cérebro. Esse enquadramento é demasiado rudimentar para ser útil. O CB1 e o CB2 são receptores acoplados à proteína G, ou GPCRs, e, tal como outros GPCRs, não actuam como simples interruptores de ligar/desligar. Traduzem sinais dos endocannabinoids produzidos pelo corpo, dos phytocannabinoids da Cannabis sativa e de ligandos sintéticos concebidos em laboratório em respostas celulares variáveis. A resposta que surge depende de onde o receptor se encontra, de qual ligando se liga, dos parceiros de sinalização disponíveis, da duração da estimulação do receptor e de o receptor ser orientado para sinalização por proteína G, recrutamento de β-arrestina, dessensibilização ou internalização.
Isto é importante porque a ciência do cannabis não diz respeito apenas à intoxicação. Diz também respeito à dor crónica, à epilepsia, à sinalização imunitária, à neurodegeneração, ao risco psiquiátrico e ao motivo pelo qual tantos programas de fármacos cannabinoid pareciam promissores em estudos pré-clínicos e depois falharam em seres humanos. As consequências são grandes. A Organização Mundial de Saúde estimou que 200 milhões de pessoas consumiram cannabis em 2019, cerca de 4% da população mundial com idades entre os 15 e os 64 anos. A epilepsia afecta cerca de 50 milhões de pessoas em todo o mundo. A esquizofrenia afecta cerca de 24 milhões. Ainda assim, em 2025, a U.S. FDA refere a aprovação de um medicamento derivado de cannabis e de três medicamentos relacionados com cannabis. Essa distância entre exposição maciça e terapêuticas aprovadas limitadas é uma das razões pelas quais a biologia dos receptores é tão relevante.
| Característica | CB1 | CB2 |
|---|---|---|
| Abreviatura típica | “recetor cerebral” | “recetor imunitário” |
| Correção do artigo | Enriquecimento central, mas também expressão periférica | Enriquecido no sistema imunitário, mas não irrelevante para o cérebro |
| Funções exemplificadas mencionadas | Perceção, memória, controlo motor, nociceção | Sinalização de citocinas, migração celular, funções neuroinflamatórias |
| Interpretação | Dependente do circuito e do estado | Dependente do tipo celular e do estado da doença |
Porque a biologia dos receptores explica mais do que as etiquetas da planta
Etiquetas como “indica”, “sativa” ou mesmo “THC-dominante” e “CBD-dominante” contam apenas parte da história, porque são os receptores, e não as categorias de marketing da planta, que estão mais próximos do mecanismo. O Δ9-tetrahydrocannabinol (THC) é um agonista parcial do CB1 e do CB2, mas o efeito a jusante do THC não é fixo. Em neurónios corticais ricos em CB1, pode suprimir a libertação de neurotransmissores e alterar a percepção, a memória e o controlo motor. Em vias sensitivas periféricas, a mesma família de receptores pode modular a nocicepção. Em células imunitárias, a activação do CB2 pode alterar a sinalização das citocinas ou a migração celular. A mesma família. Resultados diferentes.
A regra simples de que o CB1 é apenas cerebral e o CB2 é apenas imunitário é demasiado simplista para a biologia atual dos recetores.Strong evidence
A antiga regra prática — CB1 no cérebro, CB2 nas células imunitárias — baseava-se num padrão real, mas envelheceu mal. A distribuição é em gradiente e específica do tipo celular, não binária. O CB1 é fortemente expresso em muitas regiões do sistema nervoso central, especialmente em terminais pré-sinápticos, embora também apareça em tecidos periféricos. O CB2 está fortemente associado à função imunitária, mas a afirmação de que é irrelevante para o cérebro já não é defensável. Uma revisão de 2026 na Frontiers in Behavioral Neuroscience argumentou que a sinalização do CB2 tem ganho atenção em perturbações do sistema nervoso central, especificamente através de mecanismos neuroinflamatórios e neurodegenerativos, descrevendo-o como “uma actualização dos últimos 3 anos”. Essa actualização importa. Se o CB2 contribui para a patologia central em algumas condições, então os fármacos direccionados ao CB2 não podem ser compreendidos como ferramentas puramente periféricas.
A estrutura aprofunda a história. Uma revisão de 2026 na Frontiers in Chemical Biology explicou que a selectividade de ligandos entre “CB1 e CB2” resulta de diferenças estruturais ao nível do receptor, que influenciam a pose de ligação, a eficácia e a regulação do receptor. Em termos simples, pequenas alterações químicas podem enviesar um ligando para um subtipo de receptor ou para uma via de sinalização específica, o que ajuda a explicar porque é que dois cannabinoid que parecem semelhantes no papel podem sentir-se ou funcionar de forma muito diferente in vivo. Um estudo indexado no PubMed de 2025/2026 sobre selectividade de subtipos levou isto mais longe ao mostrar que a selectividade dos endocannabinoids está ligada à dinâmica conformacional do receptor, e não a um modelo rígido de chave-fechadura. O receptor move-se. O ligando estabiliza alguns estados mais do que outros. A biologia segue esses estados.
Dos phytocannabinoids à sinalização dos endocannabinoids
{kind=link}
Sinalização retrógrada Um padrão de sinalização sináptica no qual uma célula pós-sináptica liberta um mensageiro que recua para atuar sobre recetores pré-sinápticos.
Sequência de descoberta em resumo
- 1 O THC foi isolado e estruturalmente definido.
- 2 Foram demonstrados locais específicos de ligação aos canabinoides em tecido cerebral.
- 3 O CB1 foi clonado como um GPCR.
- 4 O CB2 foi identificado a partir de tecidos relacionados com o sistema imunitário.
- 5 A anandamida e depois o 2-AG estabeleceram um sistema de sinalização endógeno.
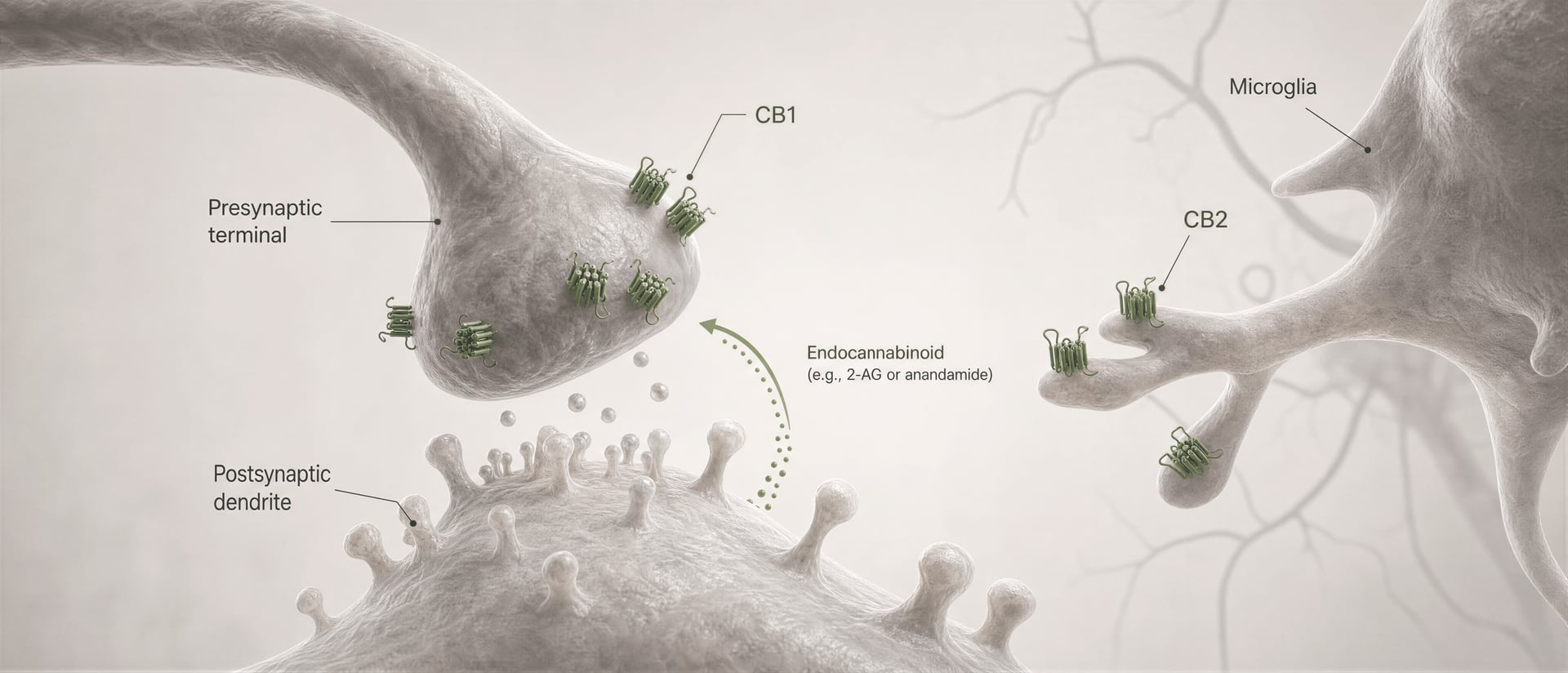
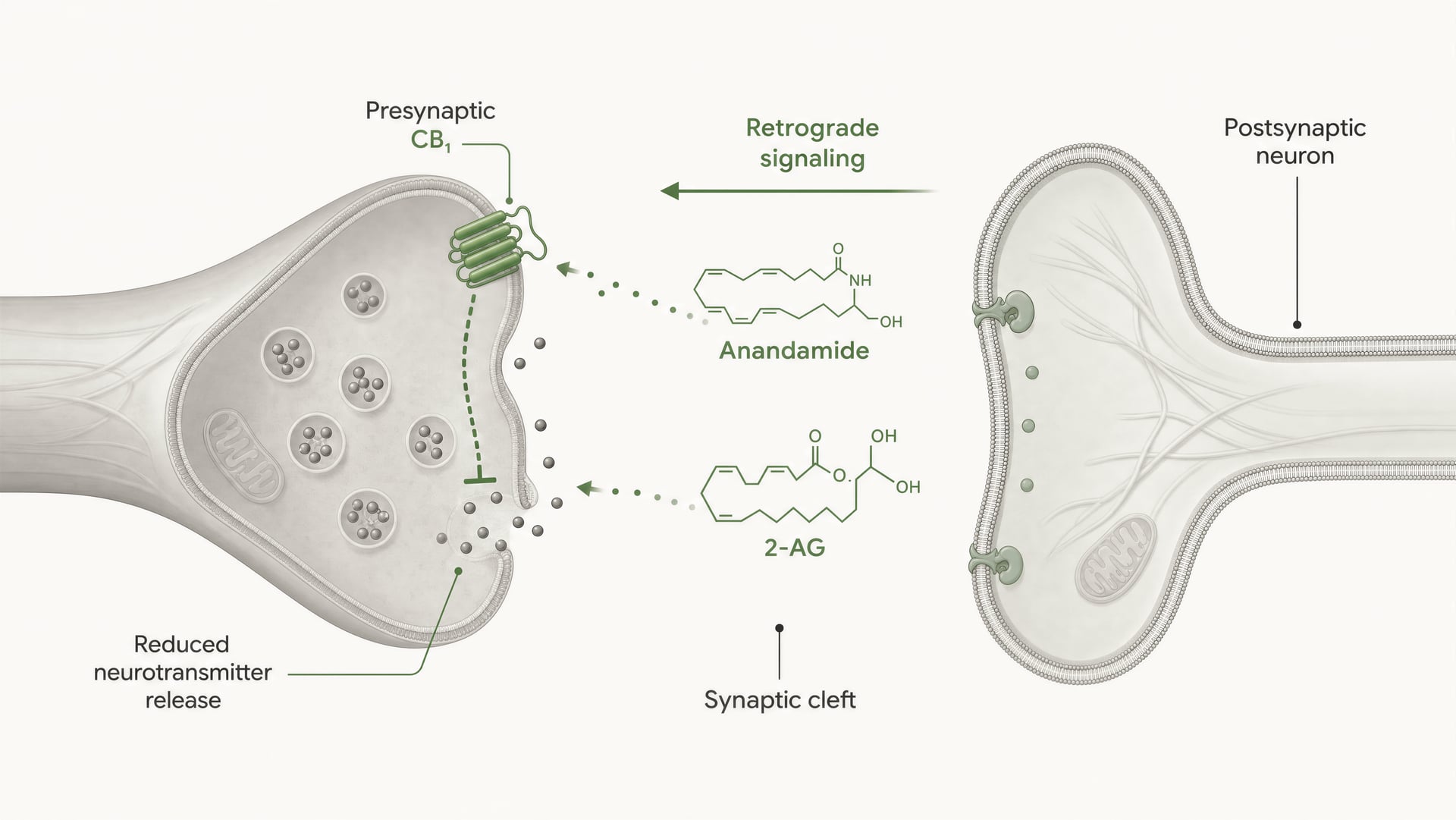
Os receptores cannabinoid não foram descobertos porque o corpo tivesse evoluído para o cannabis. A sequência foi a inversa. O trabalho de Allyn Howlett e colaboradores foi central para definir a farmacologia dos receptores cannabinoid, e a posterior descoberta da anandamida por Raphael Mechoulam e Lumír Hanuš ajudou a estabelecer que os seres humanos produzem as suas próprias moléculas de sinalização semelhantes aos cannabinoid. A anandamida e o 2-arachidonoylglycerol, habitualmente abreviado para 2-AG, são os principais endocannabinoids. São produzidos sob demanda, não armazenados em vesículas como os neurotransmissores clássicos, e frequentemente sinalizam de forma retrógrada: uma célula pós-sináptica gera um endocannabinoid que viaja para trás através da sinapse para activar o CB1 pré-sináptico e reduzir a libertação adicional de transmissores.
Isto é uma imagem fundamentalmente diferente de “substância da erva atinge o receptor”. A sinalização dos endocannabinoids é local, transitória e rigorosamente regulada por enzimas de síntese e degradação. Os phytocannabinoids entram neste sistema a partir do exterior. Os ligandos sintéticos podem atingi-lo de forma ainda mais intensa ou mais selectiva. O resultado é que o mesmo receptor pode ser activado por um impulso endógeno breve, por um phytocannabinoid oral de absorção lenta ou por um agonista sintético de elevada eficácia com responsabilidades de segurança muito diferentes.
Esta diferença é uma das razões pelas quais a intoxicação não pode ser inferida apenas pelo nome do receptor. Depende da eficácia do ligando, da dose, da via de administração, do momento e do contexto tecidular. O THC no CB1 está no centro dos efeitos psicoactivos, sim, mas esse facto não reduz o CB1 a um “receptor de psicoactividade”. Nem faz do CB2 um simples regulador anti-inflamatório. O artigo de 2025 no American Journal of Psychiatry sobre sinalização enviesada do CB1 fez precisamente este ponto mais amplo ao argumentar que ligandos enviesados para o CB1 podem oferecer uma estratégia terapêutica para a esquizofrenia. Essa proposta liga a ciência do cannabinoid a uma ideia mais ampla dos GPCRs: se um ligando favorecer ramos de sinalização benéficos enquanto evita outros associados a efeitos adversos, a acção farmacológica pode ser separada da activação bruta do receptor. Se essa promessa se confirmar clinicamente continua em aberto, mas o argumento mecanístico é sólido.
O que este artigo quer dizer com distribuição, sinalização e alvos terapêuticos
Neste artigo, distribuição significa mais do que um mapa de órgãos. Inclui densidade do receptor, tipo celular, localização subcelular, estado da doença e mudança temporal. Um receptor expresso em terminais GABAérgicos pode ter efeitos de circuito diferentes do mesmo receptor em terminais glutamatérgicos. Um receptor regulado em alta durante a inflamação não é equivalente ao seu estado basal. A distribuição é dinâmica.
Sinalização significa as consequências intracelulares do envolvimento do receptor. Para o CB1 e o CB2, isso inclui o acoplamento a proteínas G da família Gi/o, a inibição da adenilil ciclase, a modulação de canais iónicos, alterações em cascatas de cinases, recrutamento de β-arrestina, dessensibilização do receptor e internalização. Inclui também modulação alostérica e agonismo enviesado, em que os ligandos podem favorecer determinados resultados de sinalização em detrimento de outros. Isto não é um detalhe académico. É muitas vezes a diferença entre analgesia, sedação, tolerância, disfória ou um ensaio falhado.
Alvos terapêuticos significa receptores considerados para intervenção, não histórias de sucesso garantidas. O direccionamento selectivo do CB1 pode reduzir alguns efeitos off-target, mas ainda assim enfrentar eventos adversos centrais. O direccionamento selectivo do CB2 pode evitar algumas responsabilidades intoxicantes, mas a selectividade não garante eficácia em doença humana complexa. O trabalho em biologia de sistemas deixa isso claro. Uma análise integrativa de redes indexada no PubMed em 2025/2026 identificou o CB1 e o CB2 como nós altamente influentes no sistema endocannabinoid e associou a sua sinalização a vias metabólicas mais amplas. Por outras palavras, estes receptores inserem-se em redes maiores. Ao pressionar um nó, outras vias deslocam-se.
Essa é a posição deste artigo. O CB1 e o CB2 são nós de sinalização dependentes do contexto. Não são interruptores estáticos. Não são meras etiquetas para “cérebro” e “sistema imunitário”. Se a ciência do cannabis quiser explicar porque é que um composto parece terapêutico num cenário, intoxicante noutro e decepcionante numa clínica, tem de começar ao nível do receptor e permanecer aí tempo suficiente para seguir a biologia até onde ela realmente vai.
Uma breve história da descoberta dos receptores cannabinoid
Antes de os receptores cannabinoid serem identificados, a ciência do cannabis era sobretudo uma história de química. Os investigadores podiam isolar compostos da planta, comparar efeitos comportamentais brutos em animais e discutir a potência, mas ainda não conseguiam explicar como uma molécula como delta-9-tetrahydrocannabinol, ou THC, produzia os seus efeitos com qualquer coisa parecida com precisão ao nível do receptor. Isso mudou no final da década de 1980 e no início da década de 1990. A mudança foi decisiva: a investigação sobre cannabis passou de catalogar fitocannabinoids para estudar interações ligando-receptor, sinalização intracelular, distribuição tecidual e, por fim, o sistema lipídico endógeno agora chamado sistema endocannabinoid, ou ECS.
| Ano | Marco | Pessoas mencionadas no artigo |
|---|---|---|
| 1964 | Isolamento e estrutura do THC | Raphael Mechoulam; Yechiel Gaoni |
| 1988 | Locais específicos de ligação aos canabinoides com elevada afinidade em membranas do cérebro de rato | Allyn Howlett; William Devane |
| 1990 | Clonagem do CB1 | Lisa Matsuda e colaboradores |
| 1992 | Identificação da anandamida | William Devane; Lumír Hanuš; Raphael Mechoulam; colaboradores |
| 1993 | Identificação do CB2 | Sean Munro; Kerrie Thomas; M. Abu-Shaar |
| 1995 | Identificação do 2-AG por grupos separados | Equipa de Mechoulam; grupo de Sugiura |
Da farmacologia do THC à identificação do receptor
Um marco inicial importante ocorreu em 1964, quando Raphael Mechoulam e Yechiel Gaoni relataram o isolamento e a estrutura do THC. Esse feito foi relevante porque deu aos farmacologistas uma molécula definida para testar, em vez de um extrato botânico variável. Durante as duas décadas seguintes, o campo construiu um mapa estrutura-atividade a partir do THC e de outros cannabinoids relacionados, mas ainda existia debate sobre o mecanismo. Alguns investigadores favoreciam efeitos inespecíficos da membrana, porque os cannabinoids são lipofílicos. Essa posição tornou-se mais difícil de defender à medida que se acumulavam dados de ligação estereosseletiva e saturável.
A era dos receptores começou verdadeiramente com estudos de ligação na década de 1980. Em 1988, Allyn Howlett e William Devane publicaram um artigo marcante em Molecular Pharmacology mostrando sítios específicos de ligação de cannabinoids, com elevada afinidade, em membranas cerebrais de ratos, utilizando o agonista sintético CP55,940. O resultado não foi uma sugestão vaga de um alvo. Demonstrou saturabilidade, variação regional e especificidade farmacológica compatíveis com um receptor genuíno. O tecido cerebral não estava a responder aos cannabinoids como se estes se limitassem a dissolver-se nas bicamadas lipídicas e a perturbar tudo de uma vez. Havia seletividade.
Três anos mais tarde, em 1990, Lisa Matsuda e colaboradores clonaram o primeiro receptor cannabinoid, agora denominado CB1, e publicaram-no em Nature. O CB1 foi identificado como um recetor acoplado à proteína G, ou GPCR, uma descoberta que inseriu imediatamente a farmacologia cannabinoid numa das mais importantes superfamílias de sinalização da biologia. Isso foi relevante porque os GPCR não são apenas interruptores. Adotam múltiplos estados conformacionais, acoplam-se a diferentes parceiros intracelulares, dessensibilizam-se, internalizam-se e apresentam enviesamento de sinalização dependente do ligando. Essas ideias só se tornariam centrais muito mais tarde, mas a clonagem do CB1 tornou-as possíveis.
O CB2 surgiu logo de seguida. Em 1993, Sean Munro, Kerrie Thomas e M. Abu-Shaar identificaram um segundo receptor cannabinoid, o CB2, também em Nature, inicialmente caracterizado a partir de tecidos relacionados com o sistema imunitário. Esta descoberta criou uma abreviação duradoura que moldou o campo durante anos: CB1 como o “receptor cerebral” associado à intoxicação, CB2 como o receptor “periférico” ou imunitário associado à inflamação. A abreviação era útil, mas já então era demasiado grosseira, e envelheceu mal. A distribuição de ambos os receptores depende da espécie, do tipo celular, do estado de ativação, do contexto da doença e do método de ensaio.
Como o CB1 e o CB2 mudaram o campo do endocannabinoid
Depois de CB1 e CB2 terem sido identificados, a próxima pergunta óbvia foi porque é que o corpo teria receptores para cannabinoids de origem vegetal. A resposta chegou em 1992, quando William Devane, Lumír Hanuš, Raphael Mechoulam e colaboradores identificaram a anandamide, formalmente araquidonoiletanolamida, como um ligando endógeno. O artigo, publicado em Science, marcou uma rutura conceptual. A farmacologia do cannabis deixou de se centrar apenas em compostos exógenos de Cannabis sativa. Passou a tratar de um sistema nativo de sinalização lipídica.
Um segundo ligando endógeno importante, 2-arachidonoylglycerol, ou 2-AG, foi identificado em 1995 por grupos separados, incluindo a equipa de Mechoulam e o grupo de Sugiura. Com receptores e ligandos endógenos em posição, o ECS expandiu-se rapidamente. Os investigadores identificaram enzimas de síntese e degradação, como a fatty acid amide hydrolase, FAAH, para a anandamide, e a monoacylglycerol lipase, MAGL, para o 2-AG. Enfrentaram também uma questão ainda não resolvida: como é que estas moléculas altamente lipofílicas se deslocam através das membranas e do espaço extracelular. O campo fala frequentemente de “transporte”, mas permanece indescritível um transportador endocannabinoid único e dedicado.
Foi neste ponto que a ciência cannabinoid deixou de ser um esquema de dois receptores e passou a ser uma rede de sinalização. CB1 e CB2 foram ligados às proteínas Gi/o, à inibição da adenylyl cyclase, à modulação de canais de cálcio e potássio e à supressão da libertação de neurotransmissores. Mas a história não ficou tão simples. Os receptores podiam recrutar beta-arrestins, sofrer dessensibilização e internalização, e responder de forma diferente a phytocannabinoids, endocannabinoids e ligandos sintéticos, mesmo quando esses ligandos atingiam nominalmente o mesmo receptor. A linguagem atual dos GPCR de agonismo enviesado adapta-se especialmente bem aos cannabinoids. Um artigo de 2025 do American Journal of Psychiatry argumentou que a sinalização enviesada do CB1 é uma estratégia terapêutica plausível para a esquizofrenia, uma perturbação que afeta cerca de 24 milhões de pessoas em todo o mundo segundo a WHO. Isso está muito longe da antiga imagem do CB1 como meramente o receptor que explica por que motivo o THC intoxica.
A história do CB2 também se alargou. Os primeiros trabalhos colocavam-no sobretudo nos tecidos imunitários, e isso estava correto em termos gerais, mas estudos posteriores encontraram expressão de CB2 na microglia e, em algumas condições, noutras populações celulares do sistema nervoso central. Uma revisão de 2026 em Frontiers in Behavioral Neuroscience descreveu “an update over the last 3 years” ligando a sinalização de CB2 a mecanismos neuroinflamatórios e neurodegenerativos, deixando claro que o CB2 não pode ser descartado como irrelevante para o cérebro. O trabalho estrutural atual foi ainda mais longe. Uma revisão de 2026 em Frontiers in Chemical Biology sobre “CB1 and CB2” sublinhou que a seletividade de subtipo depende de diferenças estruturais ao nível do receptor que alteram a ligação, a eficácia e a regulação. Um estudo recente indexado no PubMed sobre seletividade de subtipo defende igualmente que a seletividade dos endocannabinoids é dinâmica, moldada pelo comportamento conformacional e não por um simples modelo de chave-fechadura.
Investigadores fundadores e porque é que a história ainda importa
Três nomes pertencem ao centro desta história. Raphael Mechoulam ajudou a definir a base química e biológica da ciência cannabinoid, desde o trabalho estrutural sobre THC até à descoberta de endocannabinoids. Lumír Hanuš foi uma figura central na identificação da anandamide e na investigação posterior sobre endocannabinoids. A farmacologia de receptores de Allyn Howlett foi decisiva para provar que os cannabinoids atuam através de sítios de ligação cerebrais específicos e de mecanismos de sinalização. Sem o seu trabalho, não existe o campo moderno do ECS.
A história ainda importa porque as antigas simplificações continuam a distorcer os debates atuais. Em 2019, estimava-se que 200 milhões de pessoas em todo o mundo, cerca de 4% das pessoas entre os 15 e os 64 anos, usavam cannabis segundo a WHO. Ao mesmo tempo, a FDA afirma ter aprovado um produto medicamentoso derivado de cannabis e três produtos medicinais relacionados com cannabis. A exposição pública é enorme. A tradução clínica é seletiva e difícil. A história dos receptores explica porquê. Os efeitos cannabinoid dependem da classe do ligando, do estado do receptor, da localização tecidual, do timing e do enviesamento da via. Dependem também de uma rede mais ampla. Uma análise de rede integrativa de 2025/2026 identificou CB1 e CB2 como nós altamente influentes ligados a vias metabólicas, em vez de alvos isolados.
Esse é o verdadeiro legado da descoberta dos receptores. Ela não simplificou a biologia do cannabis. Mostrou porque é que a biologia é mais complexa do que a antiga divisão entre cérebro e corpo jamais permitiu.
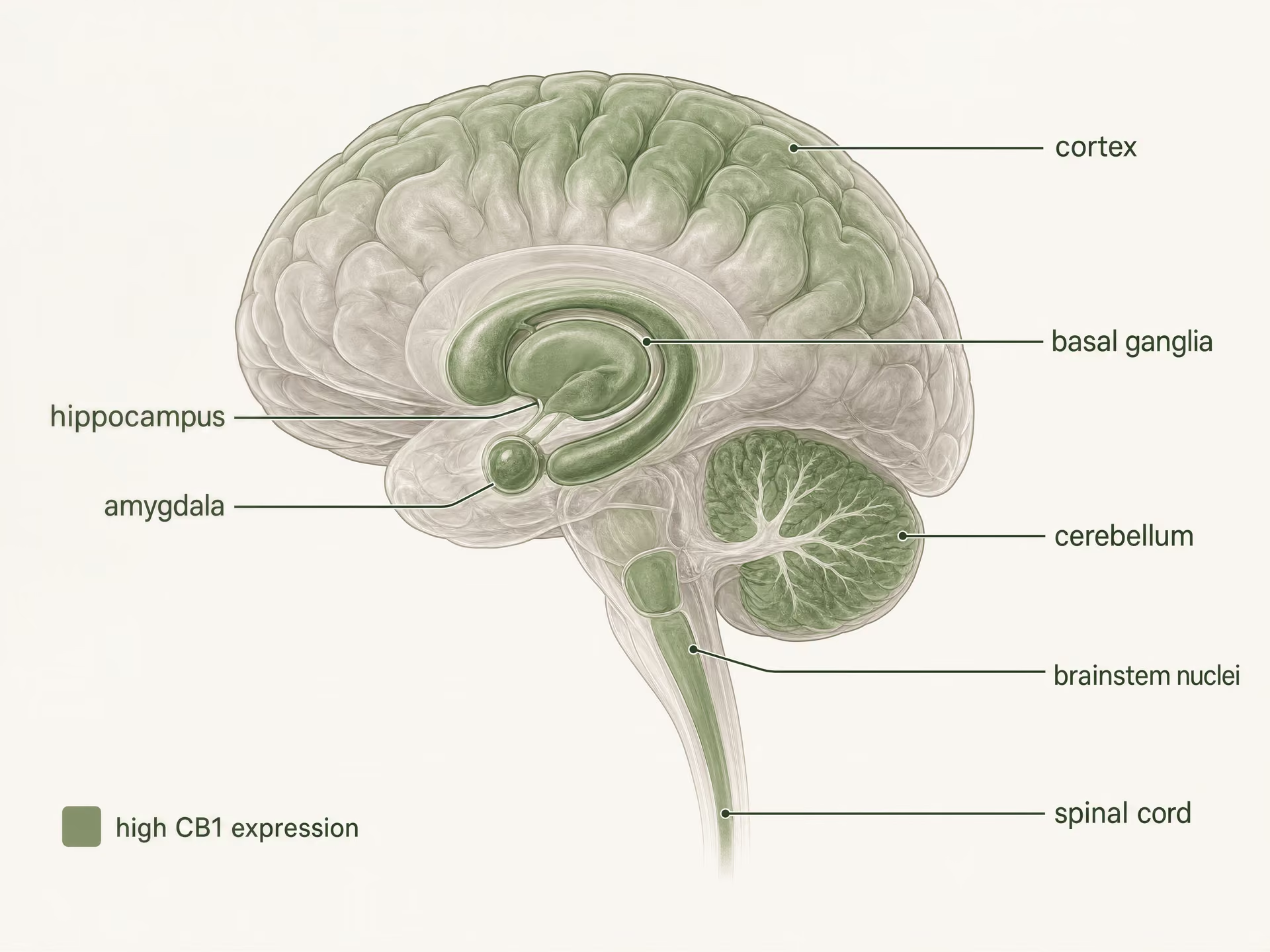
Onde se encontra o CB1: circuitos cerebrais, tecidos periféricos e gradientes funcionais
O CB1 conquistou a sua reputação de principal receptor cannabinoid psicoativo por uma razão. É abundante no sistema nervoso central, e o trabalho de farmacologia recetorial de Allyn Howlett ajudou a estabelecer que o THC atua através de um sistema recetorial específico e saturável, em vez de por efeitos inespecíficos da membrana. Mas a velha abreviação — CB1 no cérebro, CB2 nas células imunitárias — agora gera mais confusão do que clareza. O CB1 está fortemente enriquecido em circuitos neuronais, sim. Mas também está presente no intestino, fígado, tecido adiposo, órgãos reprodutores, tecidos cardiovasculares e vias sensoriais, onde molda a alimentação, o metabolismo, a sinalização da dor e a função autonómica. A distribuição é ampla. A função é condicional.
Isso importa porque a exposição a cannabinoids é comum. A World Health Organization estimou que 200 milhões de pessoas usaram cannabis em 2019, cerca de 4% da população global com idades entre 15 e 64 anos. Também importa porque a farmacologia de receptores continua a entrar na medicina: a U.S. FDA afirma que um produto medicamentoso derivado de cannabis e três produtos medicinais relacionados com cannabis estão aprovados. Um receptor encontrado em tantos órgãos não pode ser reduzido a um único rótulo comportamental.
- Padrão geral
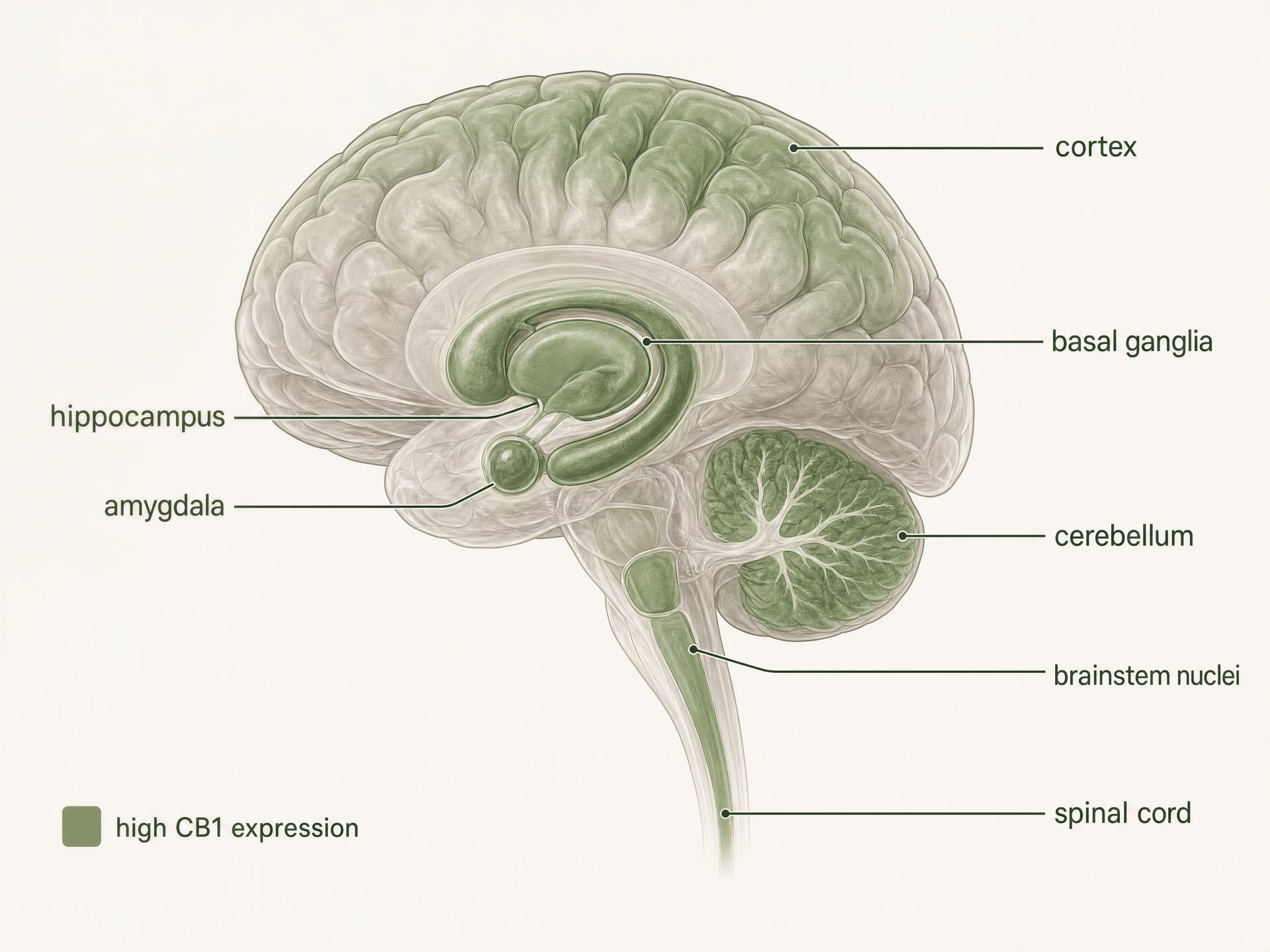
- Um dos GPCR mais abundantes no cérebro dos mamíferos
- Regiões de elevada densidade mencionadas
- Córtex, hipocampo, amígdala, gânglios da base, cerebelo
- Locais relacionados com a dor mencionados
- Substância cinzenta periaquedutal, medula ventromedial rostral, corno dorsal
- Locais periféricos mencionados
- Intestino, fígado, tecido adiposo, vias reprodutivas, cardiovasculares e sensoriais
Elevada expressão no sistema nervoso central
{kind=link}
O CB1 é um dos recetores acoplados à proteína G mais abundantes no cérebro dos mamíferos. A autoradiografia, a hibridação in situ e o mapeamento imunohistoquímico construíram uma imagem clara muito antes dos estudos estruturais atuais: surgem densidades elevadas no córtex, hipocampo, amígdala, gânglios da base, cerebelo e várias regiões de processamento da dor, com expressão adicional nos núcleos do tronco cerebral e ao longo da medula espinal. Este padrão corresponde aos efeitos clássicos do THC de forma surpreendentemente boa, mas não perfeita.
| Localização do CB1 | Efeito sináptico imediato | Consequência exemplificada mencionada no artigo |
|---|---|---|
| Terminais de interneurónios GABAérgicos | Suprime a libertação de GABA | Desinibição dos neurónios a jusante |
| Terminais glutamatérgicos | Suprime a libertação de glutamato | Excitação atenuada |
| Circuitos dos gânglios da base e do cerebelo | Altera a libertação de neurotransmissores nas vias motoras | Lentificação motora, alteração dos circuitos de hábito, coordenação comprometida |
| Vias da dor | Modula a transmissão nociceptiva | Alterações no processamento ascendente, descendente, inflamatório e afetivo da dor |
No córtex e no hipocampo, o CB1 encontra-se em circuitos que regulam a atenção, a memória de trabalho, a aprendizagem da extinção e a plasticidade sináptica a curto prazo. Os efeitos na memória não se resumem a “hipocampo=esquecimento”. Dependem fortemente de quais terminais axonais exprimem o receptor. O CB1 está frequentemente concentrado na região pré-sináptica, onde suprime a libertação de neurotransmissores após ativação por endocannabinoid tais como a anandamida e o 2-arachidonoylglycerol, os lípidos de sinalização cuja descoberta por Raphael Mechoulam, Lumír Hanuš e colaboradores transformou o campo. Quando o CB1 é ativado nos terminais de interneurónios GABAérgicos, pode desinibir neurónios a jusante; quando é ativado nos terminais glutamatérgicos, pode atenuar a excitação. O mesmo receptor, resultado de rede oposto.
Os gânglios da base e o cerebelo explicam outro conjunto familiar de efeitos. A expressão densa de CB1 no estriado, no globo pálido, na substância negra pars reticulata e nas camadas moleculares do cerebelo associa a ativação do receptor a lentificação motora, alteração dos circuitos do hábito, coordenação prejudicada e, em algumas doses, efeitos do tipo catalepsia em modelos animais. Contudo, o facto de o CB1 ser escasso nos centros cardiorrespiratórios do tronco cerebral, em comparação com os receptores opioides, ajuda a explicar por que razão a sobredosagem de cannabinoid não produz, em geral, o mesmo padrão de depressão respiratória fatal observado com agonistas opioides potentes. A localização importa. O que está ausente também.[5]The Health Effects of Cannabis and Cannabinoids: The Current State of Evidence and Recommendations for Research. National Academies of Sciences, Engineering, and Medicine. National Academies Press, 2017. https://nap.nationalacademies.org/catalog/24625/the-health-effects-of-cannabis-and-cannabinoids-the-current-state
O processamento da dor mostra a mesma lógica regional. O CB1 é encontrado na substância cinzenta periaquedutal, na medula rostroventromedial, no corno dorsal da medula espinal e nas vias nociceptivas periféricas. Isso confere ao receptor múltiplos pontos de entrada na nociceção: pode alterar os sinais ascendentes de dor, o controlo descendente da dor, a sensibilização inflamatória e a coloração emocional da dor. Esta é uma das razões pelas quais os cannabinoid permanecem na discussão da dor crónica, sobretudo porque quase 1 em cada 5 adultos nos Estados Unidos vive com dor crónica, segundo as National Academies. Mas a analgesia não está garantida apenas porque o CB1 está presente. Sedação, prejuízo cognitivo, tolerância e efeitos adversos limitantes da dose surgem frequentemente através de circuitos adjacentes ou dos mesmos circuitos em diferentes níveis de ativação do receptor.
A sinalização enviesada do CB1 pode separar efeitos terapêuticos desejados de efeitos psicoativos ou cognitivos indesejados.Limited evidence
Proteínas Gi/o Uma família de proteínas G que, em geral, reduz a atividade da adenilil ciclase e ajuda a controlar canais iónicos após a ativação do GPCR.
A biologia moderna dos recetores acrescenta outra camada. O CB1 não é um simples interruptor de ligar/desligar. Acopla-se sobretudo a proteínas Gi/o, reduzindo a atividade da adenilil ciclase e modulando canais iónicos, mas também pode recrutar beta-arrestinas, sofrer dessensibilização e internalização, e apresentar enviesamento de sinalização dependente do ligando. O artigo de 2025 do American Journal of Psychiatry que argumenta que a sinalização enviesada de CB1 poderia ser explorada terapeuticamente na esquizofrenia aponta isto de forma direta: a ocupação do recetor, por si só, é um fraco preditor do desfecho. Com a esquizofrenia a afetar cerca de 24 milhões de pessoas em todo o mundo, é evidente o interesse em separar a sinalização desejada dos efeitos psicoativos ou cognitivos indesejados. Se essa separação é exequível na prática continua a ser uma questão aberta de desenvolvimento de fármacos, não um facto estabelecido.
CB1 periférico no intestino, fígado, tecido adiposo e além
O CB1 fora do cérebro não é uma nota de rodapé. É central para explicar por que razão os cannabinoid afetam o apetite, a náusea, o controlo da glicose, o metabolismo lipídico e a sensação visceral.
No intestino, o CB1 é expresso no sistema nervoso entérico, em compartimentos epiteliais e em vias relacionadas com o nervo vago. A sua ativação pode retardar o esvaziamento gástrico, alterar a motilidade intestinal, reduzir o vómito e modificar a sinalização entre o intestino e o cérebro. Os efeitos sobre o apetite são muitas vezes descritos como se se originassem exclusivamente nos centros hipotalâmicos de recompensa e alimentação, mas o CB1 periférico contribui para essa história ao moldar as entradas sensoriais e hormonais antes de os sinais sequer chegarem a esses circuitos. Uma refeição não atua sobre uma paisagem recetorial em branco; altera localmente o tónus endocannabinoid.
No fígado e no tecido adiposo, o CB1 participa na regulação metabólica, incluindo lipogénese, sensibilidade à insulina e armazenamento energético. Esta foi uma das principais lições da era do rimonabant. O bloqueio do CB1 melhorou o peso e os marcadores metabólicos, apoiando a ideia de que a sinalização endocannabinoid hiperativa contribui para a patologia relacionada com a obesidade. Mas o rimonabant, um inverso agonista de CB1 com ação central, também produziu efeitos adversos psiquiátricos graves, incluindo depressão e ansiedade, e foi retirado do mercado. Esse episódio é frequentemente citado como uma falha do “alvo CB1”. Mais corretamente, foi uma falha de um tipo particular de abordagem ao CB1: antagonismo central forte ou inverso agonismo num sistema recetorial inserido em circuitos de humor e stress. A lição não é que o CB1 periférico seja irrelevante; é que o padrão de exposição ao fármaco e o estado do recetor importam tanto como o nome do recetor.
Adipócitos, hepatócitos, tecido pancreático, músculo esquelético, tecidos cardiovasculares e órgãos reprodutivos contribuem todos para o mapa periférico. O mesmo acontece com neurónios sensoriais. A análise de rede integrativa indexada no PubMed de 2025/2026 que identificou CB1 e CB2 como nós altamente influentes na sinalização endocannabinoid é útil aqui porque desloca o enquadramento da localização do recetor para a sua participação em redes metabólicas e de sinalização. Um recetor com expressão modesta num tecido pode ainda assim exercer efeitos sistémicos amplos se ocupar um ponto de estrangulamento na sinalização local.
O trabalho estrutural também mantém esta discussão rigorosa. A revisão de 2026 em Frontiers in Chemical Biology sobre CB1 e CB2 sublinha que a seletividade e a eficácia dos ligandos resultam de diferenças estruturais ao nível do recetor que alteram a ligação, a sinalização e a regulação do recetor. Um estudo indexado no PubMed de 2025/2026 sobre seletividade de subtipo defende igualmente que a dinâmica conformacional, e não apenas o encaixe chave-fechadura, molda a forma como os endocannabinoid distinguem subtipos de recetor. Isto é importante para a distribuição de CB1 porque “CB1 no fígado” não significa que o THC, a anandamida, o 2-AG e um agonista sintético façam todos a mesma coisa aí.
Porque a distribuição não equivale a uma função uniforme
O maior erro nos mapas de recetores é tratar a expressão como destino. Não é. A expressão elevada diz-lhe onde procurar, não o que vai acontecer.
Primeiro, o tipo celular altera o sinal do efeito. Um recetor CB1 num terminal glutamatérgico pode reduzir a excitação. O mesmo recetor num terminal GABAérgico pode reduzir a inibição. Esses resultados não são intercambiáveis. Segundo, a localização sináptica importa. O CB1 está normalmente na região pré-sináptica, muitas vezes ativado por endocannabinoid libertados “sob demanda” pelos neurónios pós-sinápticos, criando controlo retrógrado sobre a libertação de neurotransmissores. Essa organização favorece uma modulação breve e dependente da atividade, em vez de uma ativação constante do recetor.
Terceiro, a identidade do ligando importa. Os endocannabinoid são mensageiros locais de curta duração. Os fitocannabinoid como o THC chegam de fora do sistema, muitas vezes com exposições mais elevadas e mais duradouras do que os sinais endógenos. Os ligandos sintéticos podem ir ainda mais longe, com eficácia e enviesamento diferentes. Alguns promovem a sinalização Gi/o mais fortemente; outros favorecem o recrutamento de beta-arrestina, a dessensibilização ou a internalização do recetor. É por isso que dois compostos podem ambos ser chamados agonistas de CB1 e ainda assim diferirem acentuadamente na estimulação do apetite, na perturbação da memória, no défice motor e no desenvolvimento de tolerância.
Quarto, a disponibilidade local do ligando muda tudo. A anandamida e o 2-AG são produzidos e degradados no local, pelo que os seus efeitos dependem da atividade neuronal, do estado metabólico, da expressão enzimática e do contexto inflamatório. Quinto, a própria densidade do recetor existe num gradiente. Região cerebral, estádio de desenvolvimento, estado de doença e exposição repetida ao fármaco alteram todos os níveis de CB1 e a responsividade.
A literatura atual está a afastar-se dos binarismos precisamente por esta razão. A revisão de 2026 em Frontiers in Behavioral Neuroscience observa uma atualização nos últimos 3 anos na forma como a sinalização dos recetores cannabinoid é compreendida nos distúrbios do SNC, sobretudo quando os mecanismos neuroinflamatórios e neurodegenerativos são incluídos. O CB1 deve ser lido com a mesma cautela. É um recetor central dominante, mas não exclusivamente central; um recetor da alimentação, mas não apenas isso; um alvo para a dor, mas não um interruptor analgésico limpo. Qualquer descrição séria da distribuição de CB1 tem de pensar em gradientes, circuitos e estados de sinalização, e não numa caricatura cérebro-versus-corpo.
Onde o CB2 é encontrado: raízes no sistema imunitário e o mapa em expansão do SNC
A antiga formulação dizia-o de forma simples: o CB1 é o recetor do cérebro, o CB2 é o recetor imunitário. Esse enquadramento ajudou no ensino inicial, mas agora induz mais erro do que clareza. O CB2 mostra, de facto, um enriquecimento clássico fora dos neurónios, sobretudo em linhagens imunitárias e hematopoiéticas, e esse facto continua a ser importante para a farmacologia. No entanto, a literatura mais recente, em especial a revisão de 2026 em Frontiers in Behavioral Neuroscience, defende algo mais forte: o CB2 está agora a ser discutido em distúrbios do sistema nervoso central porque a sua expressão e sinalização se tornam mais visíveis na microglia, em circuitos inflamatórios e em estados relacionados com lesão, uma “atualização nos últimos 3 anos” que alterou a forma como o recetor é mapeado e interpretado. O resultado não é que o CB2 se tenha subitamente tornado um recetor cerebral pan-neuronal, de elevada abundância. Não se tornou. O resultado é que a distribuição do recetor tem de ser descrita como condicional, específica do tipo celular e dependente do estado.
Essa distinção é clinicamente relevante. A Organização Mundial da Saúde estimou que 200 milhões de pessoas consumiram cannabis em 2019, ou 4% da população mundial com idades entre os 15 e os 64 anos. Mesmo com apenas um pequeno número de medicamentos relacionados com cannabinoid aprovados — a FDA, em 2025, contava um produto derivado de cannabis e três produtos relacionados com cannabis — a localização dos recetores continua a orientar onde os promotores de fármacos procuram efeitos anti-inflamatórios, analgésicos, neuroprotetores e psiquiátricos, e onde esperam efeitos adversos.
- Enriquecimento clássico
- Células imunitárias e hematopoiéticas
- Tipos celulares mencionados
- Linfócitos B, linfócitos T, macrófagos, monócitos, células natural killer, neutrófilos, mastócitos
- Tecidos canónicos mencionados
- Baço, amígdala palatina, timo, medula óssea, células imunitárias circulantes
- Relevância para o SNC enfatizada
- Microglia e estados ligados à patologia
Enriquecimento clássico em células imunitárias e hematopoiéticas
CB2 foi originalmente identificado como o subtipo de recetor de cannabinoid com maior expressão em células ligadas à imunidade, e não à transmissão sináptica rápida. Esse continua a ser o ponto de partida correto. Em comparação com CB1, que está amplamente প্রতিনিধado em muitas populações neuronais, CB2 é classicamente enriquecido em células B, células T, macrófagos, monócitos, células natural killer, neutrófilos, mastócitos e outros compartimentos hematopoiéticos. Assim, o baço, as amígdalas, o timo, a medula óssea e as populações circulantes de células imunitárias têm sido os tecidos canónicos para a análise de CB2.
Esta ênfase imunitária moldou a ideia inicial de desenvolvimento de fármacos de agonistas seletivos de CB2 como forma de obter benefícios anti-inflamatórios ou analgésicos, evitando simultaneamente os efeitos intoxicantes associados à forte ativação de CB1 no cérebro. Era uma hipótese sensata, mas apenas parcialmente completa. CB2 é um GPCR acoplado a Gi/o e, tal como CB1, não funciona simplesmente como um interruptor “ligado” ou “desligado”. Dependendo do ligando, da conformação do recetor e do contexto celular, CB2 pode reduzir a atividade da adenilil ciclase, influenciar vias MAPK, alterar indiretamente o acoplamento de canais iónicos, recrutar beta-arrestinas e sofrer dessensibilização ou internalização. Por isso, mesmo em tecidos imunitários periféricos, a verdadeira questão não é apenas se CB2 está presente, mas em que células é expresso, em que nível, sob que estímulo e com que enviesamento a jusante.
Essa complexidade é uma das razões pelas quais ligandos com aspeto semelhante podem comportar-se de forma diferente. A revisão de 2026 na Frontiers in Chemical Biology sobre a estrutura dos recetores de cannabinoid defende que a seletividade em “CB1 e CB2” é moldada por diferenças estruturais ao nível do recetor, que alteram a ligação do ligando, a eficácia da sinalização e a regulação do recetor. Um estudo indexado no PubMed de 2025/2026 sobre seletividade de subtipos levou o argumento mais longe ao mostrar que a seletividade dos endocannabinoid é dinâmica e está ligada a estados conformacionais, e não a um modelo rígido de chave-fechadura. Isso é importante para o mapeamento tecidual, porque um ligando endógeno como o 2-AG ou a anandamida, um phytocannabinoid como o THC, e um agonista sintético com preferência para CB2 podem todos encontrar a mesma população de recetores, mas estabilizar resultados de sinalização diferentes.
Assim, o mapa antigo de CB2 centrado no sistema imunitário não estava errado. Estava incompleto. CB2 continua a ser melhor descrito como um recetor com fortes raízes no sistema imunitário. Mas as raízes não são o organismo inteiro.
| Contexto | Como o CB2 é descrito | Ponto interpretativo |
|---|---|---|
| Cérebro saudável de base | Muitas vezes baixo ou próximo dos limites de deteção em अनेक regiões | Sinal basal baixo não equivale a irrelevância |
| Microglia ativada | Mais detetável após lesão ou inflamação | Sustenta a relevância no SNC através de funções semelhantes às imunitárias |
| Astrócitos / endotélio / células infiltrantes | Relatados em alguns contextos patológicos | A localização depende do método e do modelo |
| Expressão neuronal constitutiva ampla | Requer provas mais fortes | O artigo trata esta afirmação com cautela |
CB2 em microglia, neuroinflamação e estados de lesão
O argumento mais forte para a relevância central não resulta de afirmar que CB2 é amplamente abundante em neurónios saudáveis do prosencéfalo. Resulta da microglia e da biologia da doença.
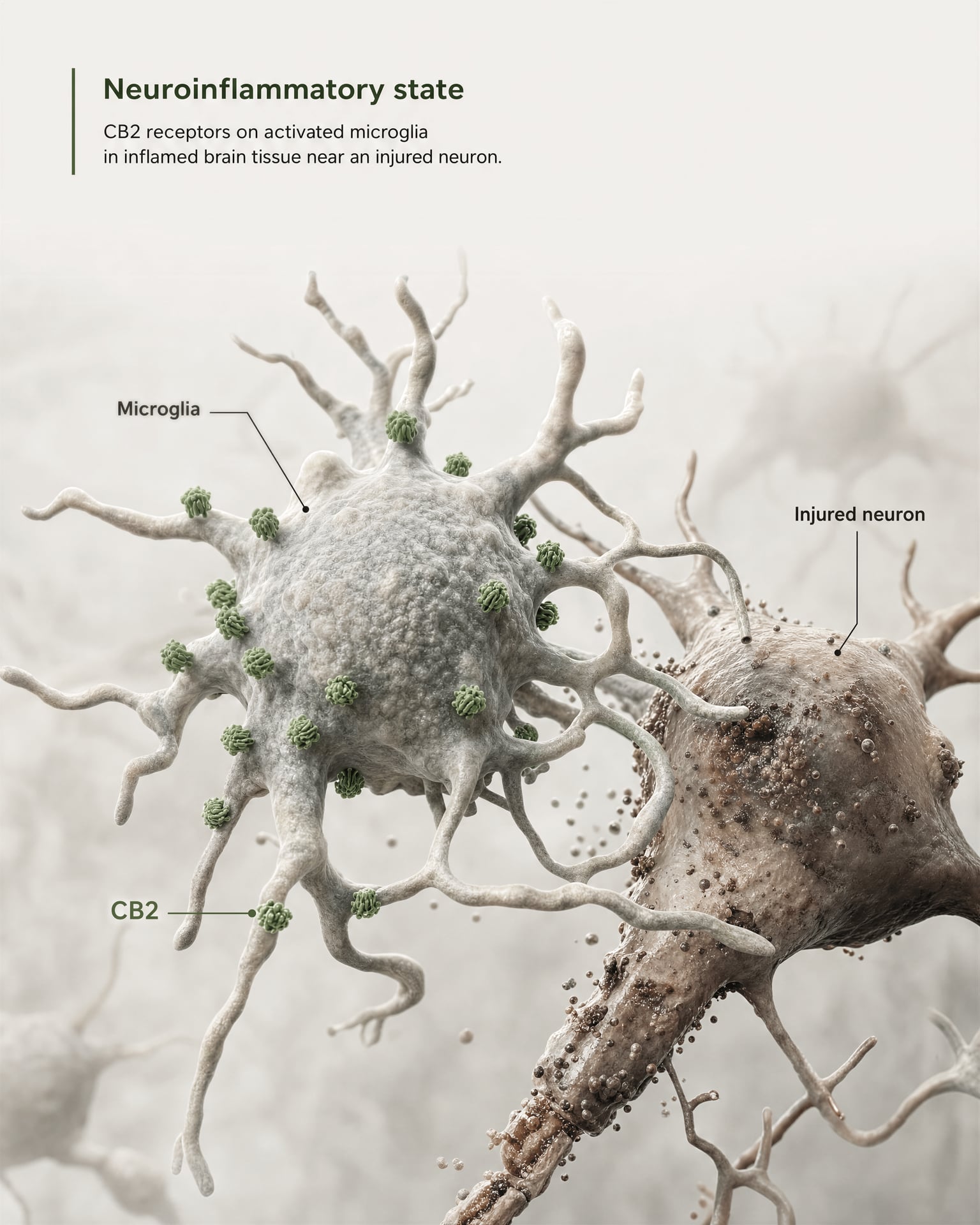
{kind=link}
A microglia são as células imunitárias residentes do SNC e situam-se exatamente na fronteira onde o antigo modelo de “recetor imunitário periférico” começa a falhar. Se um recetor é expresso no próprio sistema de vigilância imunitária e na resposta inflamatória do cérebro, então chamá-lo apenas de periférico torna-se impreciso. A revisão de 2026 na Frontiers in Behavioral Neuroscience afirma isto de forma direta: a sinalização de CB2 tem recebido atenção em perturbações do SNC porque se relaciona com mecanismos neuroinflamatórios e neurodegenerativos. É por isso que CB2 surge agora em discussões sobre doença de Alzheimer, doença de Parkinson, esclerose múltipla, traumatismo cranioencefálico, AVC, dor neuropática e algumas condições psiquiátricas em que a sinalização inflamatória faz parte da patologia.
A expressão-chave é expressão induzida ou aumentada. Em muitas regiões cerebrais saudáveis, a expressão basal de CB2 é baixa, por vezes próxima dos limites dos métodos de deteção mais antigos. Mas após lesão, infeção, inflamação crónica ou neurodegeneração, o sinal de CB2 torna-se frequentemente mais detetável, sobretudo em microglia ativadas e, em alguns estudos, em astrócitos, células imunitárias infiltrantes, compartimentos endoteliais ou subconjuntos neuronais restritos. Esta é uma regra de distribuição muito diferente da aplicada habitualmente a CB1. CB1 é muitas vezes constitutivamente abundante em circuitos neuronais definidos. CB2 é mais frequentemente interpretado como um recetor cuja relevância no SNC emerge sob stress, patologia ou ativação inflamatória.
Essa distinção tem consequências práticas. Um fármaco dirigido a CB2 pode ter pouco efeito num tecido saudável onde a densidade do recetor é baixa, mas mostrar atividade mensurável num tecido doente onde a expressão aumentou e as redes de sinalização mudaram. Esta indutibilidade é uma das razões pelas quais os resultados pré-clínicos têm sido simultaneamente entusiasmantes e difíceis de transpor. O momento temporal importa. O estádio da doença importa. A composição celular importa. Um ambiente microglial pós-lesão não é farmacologicamente equivalente a uma fatia cerebral não estimulada.
Os problemas de interpretação não são triviais. CB2 tem uma longa história de preocupações com a especificidade de anticorpos, deteção de transcritos em baixa abundância, diferenças entre espécies e alegações inconsistentes de localização entre métodos. Alguns relatos iniciais terão provavelmente sobrestimado CB2 neuronal porque as ferramentas disponíveis eram fracas. Por isso, os estudos cuidadosos recorrem agora a evidência convergente—transcriptómica de célula única, hibridização in situ, repórteres genéticos validados, controlos de knockout, dados proteómicos quando possível e comparações dependentes do estado—em vez de um único resultado de coloração. Se um estudo relata CB2 em neurónios em condições basais e outro não o deteta, a discrepância pode refletir diferenças regionais reais, estado da doença, espécie, idade ou simplesmente limitações do ensaio.
O CB2 tem relevância significativa para o SNC, especialmente em contextos gliais e inflamatórios ligados a lesão.Limited evidence
Assim, a melhor posição atual é contida, mas clara: CB2 tem relevância real no SNC, sobretudo através de funções gliais e semelhantes às imunitárias, e essa relevância aumenta durante a neuroinflamação e a lesão. Alegações de expressão neuronal constitutiva ampla de CB2 no cérebro normal exigem provas mais fortes do que as alegações de CB2 microglial e associada à patologia.
Como os últimos 3 anos mudaram a conversa sobre CB2
A revisão de 2026 na Frontiers in Behavioral Neuroscience enquadra explicitamente a literatura recente como uma “atualização dos últimos 3 anos”, e essa formulação capta uma mudança real. A conversa deixou de ser sobre se CB2 está “no cérebro ou não” e passou a ser sobre onde, quando e em que estados patológicos a sua sinalização se torna acionável.
Três desenvolvimentos impulsionaram essa mudança. Primeiro, os métodos com resolução celular melhoraram. Conjuntos de dados de RNA de célula única e de núcleo único, melhor mapeamento espacial e padrões de validação mais rigorosos reduziram a probabilidade de que a expressão de baixo nível ou induzível fosse descartada simplesmente porque os ensaios antigos careciam de sensibilidade. Segundo, a neuroinflamação tornou-se central para muitos modelos de perturbações cerebrais. Uma vez que as doenças foram analisadas através de mecanismos imunitários e gliais, em vez de apenas através de quadros centrados nos neurónios, CB2 tornou-se muito mais difícil de ignorar. Terceiro, a farmacologia dos recetores amadureceu. O campo pensa agora mais em termos de eficácia, enviesamento da sinalização, tráfego do recetor e respostas dependentes do contexto do que em simples ocupação.
Essa perspetiva mais ampla sobre GPCR é visível mesmo fora da literatura sobre CB2. O artigo de 2025 no American Journal of Psychiatry sobre sinalização enviesada de CB1 e esquizofrenia defende que a farmacologia de cannabinoid deve ser entendida através da sinalização enviesada, e não por uma ativação rudimentar do recetor. A esquizofrenia afeta cerca de 24 milhões de pessoas em todo o mundo, segundo a WHO, pelo que isto não é uma questão académica lateral. A mesma lógica aplica-se a CB2. Um ligando “seletivo para CB2” no papel pode ainda assim produzir resultados diferentes consoante conduza preferencialmente a sinalização por G-proteína, o recrutamento de beta-arrestina, a internalização do recetor ou programas transcricionais anti-inflamatórios em microglia ativadas.
A visão sistémica mais recente reforça isto. Um estudo de análise de redes indexado no PubMed de 2025/2026 identificou CB1 e CB2 como nós altamente influentes no sistema endocannabinoid e ligou a sinalização dos recetores a vias metabólicas, em vez de isolar os recetores do restante da biologia celular. Isso é coerente com o que os dados de CB2 no SNC mostram: a distribuição não é uma entrada fixa de atlas. É parte de uma rede de sinalização adaptativa.
A conclusão é simples. CB2 deve continuar a ser apresentado como um recetor de cannabinoid enriquecido no sistema imunitário. Mas parar aí agora dá uma imagem errada. No cérebro, CB2 é melhor compreendido como um recetor com baixo nível basal, indutível e associado à doença, cuja importância se torna mais clara em microglia e em estados neuroinflamatórios — e cuja deteção ainda depende fortemente do método, do modelo e do momento.
Como os receptores cannabinoidais transmitem sinal: acoplamento Gi/o, segundos mensageiros e efeitos sinápticos
A farmacologia dos receptores cannabinoidais começa com uma afirmação simples que rapidamente se torna complexa: CB1 e CB2 são receptores acoplados à proteína G da classe A, e ambos transmitem sinal, na maioria das vezes, através de proteínas Gi/o. Esse facto básico, estabelecido por trabalhos fundamentais sobre receptores por Allyn Howlett e outros, mantém-se válido. O que mudou foi a compreensão do que o acoplamento Gi/o realmente significa nas células. Não significa um único efeito a jusante. Significa um conjunto de efeitos possíveis, cuja combinação depende do ligando, da densidade do recetor, do estado de fosforilação, do ambiente da membrana, do tipo celular e do tempo.
Essa distinção é importante porque cerca de 200 milhões de pessoas usaram cannabis em 2019, ou 4% da população mundial entre os 15 e os 64 anos, segundo a World Health Organization, enquanto a FDA afirma que, em 2025, estão aprovados um medicamento derivado de cannabis e três medicamentos relacionados com cannabis. A sinalização dos recetores não é um tema secundário. É o mecanismo que separa um fármaco útil contra as crises epilépticas de uma sedação, um fármaco para o apetite que falhou de efeitos adversos psiquiátricos e um ligando seletivo em laboratório de um composto clinicamente dececionante.
Sinalização canónica de GPCR em CB1 e CB2
Sequência de sinalização canónica do CB1/CB2
- Ligação do ligando Um agonista estabiliza uma conformação ativa do recetor.
- Ativação da proteína G O recetor promove a troca de GDP por GTP em Gi/o.
- Separação das subunidades Galpha e Gbeta-gamma regulam efetores a jusante.
- Alteração do segundo mensageiro A atividade da adenilil ciclase diminui e o cAMP reduz-se.
- Efeito celular Canais iónicos, libertação de neurotransmissores, cinases e regulação génica alteram-se.
No esquema canónico, a ligação do agonista estabiliza uma conformação ativa do recetor, o recetor atua como um fator de troca de nucleótidos de guanina para Gi/o, a Gαi/o troca GDP por GTP e, em seguida, os componentes Gα e Gβγ regulam efetores a jusante. Para CB1 e CB2, a leitura clássica é a inibição da adenilil ciclase e a diminuição do AMP cíclico intracelular. Esse achado tornou-se uma das primeiras assinaturas bioquímicas usadas para definir a atividade dos recetores cannabinoidais.
Mas “canónico” não deve ser lido como “uniforme”. CB1 mostra elevada atividade constitutiva em vários sistemas de expressão, o que significa que o recetor pode transmitir sinal de forma mensurável mesmo sem agonista presente. Essa propriedade ajuda a explicar por que os agonistas inversos, como o rimonabant, fizeram mais do que bloquear o tónus cannabinoidal endógeno; eles empurraram a sinalização para abaixo da linha de base e produziram efeitos adversos centrais marcados. CB2 também se acopla a Gi/o, mas a forma como os ligandos estabilizam estados ativos difere de CB1. Trabalhos estruturais revistos em Frontiers in Chemical Biology em 2026 enfatizaram que a seletividade entre os subtipos “CB1 e CB2” é determinada por diferenças ao nível do recetor que alteram não só a ligação, mas também a eficácia e a regulação. Um estudo indexado no PubMed de 2025/2026 sobre seletividade de subtipos foi ainda mais longe, argumentando que a seletividade dos endocannabinoids é dinâmica, moldada pelo comportamento conformacional e não por um modelo fixo de chave-fechadura.
Esta é uma das razões pelas quais fitocannabinoids, endocannabinoids e ligandos sintéticos nunca devem ser tratados como intercambiáveis. A anandamida, identificada por Raphael Mechoulam e Lumír Hanuš, e o 2-arachidonoylglycerol são ligandos endógenos produzidos sob demanda e rapidamente inativados. Δ9-THC é um agonista parcial de origem vegetal, com cinética e eficácia distintas das desses endocannabinoids. Agonistas sintéticos como CP55,940, WIN55,212-2 ou HU-210 frequentemente induzem uma ativação mais intensa do recetor e podem recrutar vias de sinalização em graus diferentes. Alguns ligandos favorecem a sinalização por proteína G em detrimento do recrutamento de β-arrestina; outros não. O artigo do American Journal of Psychiatry de 2025 afirmou isso de forma direta para CB1, argumentando que a sinalização enviesada é uma estratégia terapêutica plausível na esquizofrenia, uma perturbação que afeta cerca de 24 milhões de pessoas em todo o mundo.
CB2 acrescenta outra correção às simplificações antigas. Continua enriquecido em muitas populações de células imunitárias, mas a revisão de 2026 em Frontiers in Behavioral Neuroscience descreveu “uma atualização dos últimos 3 anos” na qual a sinalização de CB2 ganhou atenção em perturbações do sistema nervoso central associadas à neuroinflamação e à neurodegeneração. Assim, mesmo antes de entrar em jogo o agonismo enviesado, a antiga dicotomia entre um “recetor do cérebro” e um “recetor imunitário” já falha ao nível do contexto de sinalização.
Efeitos sobre cAMP, canais iónicos e libertação de neurotransmissores
O efeito de segundo mensageiro mais conhecido em ambos os recetores é a supressão da formação de cAMP através da inibição da adenilil ciclase. Menos cAMP significa frequentemente menor atividade da protein kinase A, fosforilação alterada de alvos a jusante e mudanças mais lentas na expressão génica através de vias como a CREB. Nos neurónios, porém, os efeitos rápidos são muitas vezes mais importantes do que os lentos.
CB1 está fortemente posicionado para o controlo pré-sináptico. Em muitos circuitos cerebrais, localiza-se em terminais axonais, onde a ativação do recetor reduz a probabilidade de libertação de neurotransmissores. Isso ocorre através de uma combinação de inibição mediada por Gβγ de canais de cálcio dependentes de voltagem e de ativação de condutâncias de potássio retificadoras de entrada ou de outras correntes de potássio que amortecem a excitabilidade do terminal. Menor entrada de cálcio significa que menos vesículas fundem-se. O resultado é menor libertação de neurotransmissor para a fenda sináptica.
Esse é o mecanismo central da sinalização retrógrada de endocannabinoids a curta distância. Um neurónio pós-sináptico torna-se ativo, sintetiza endocannabinoids sob demanda a partir de precursores lipídicos de membrana e envia-os de volta através da sinapse para ativar recetores CB1 pré-sinápticos. O terminal pré-sináptico passa então a libertar menos neurotransmissor. É um travão de feedback. Em sinapses excitatórias, isso pode suprimir a libertação de glutamato; em sinapses inibitórias, pode suprimir a libertação de GABA. A direção do output do circuito depende de qual terminal expressa CB1. Mesmo recetor, consequência de rede oposta.
| Termo | O que é suprimido | Mecanismo descrito |
|---|---|---|
| DSI | Inibição | A atividade pós-sináptica liberta endocanabinoides que ativam CB1 pré-sináptico e reduzem a libertação de GABA |
| DSE | Excitação | A atividade pós-sináptica liberta endocanabinoides que ativam CB1 pré-sináptico e reduzem a libertação de glutamato |
DSI e DSE Formas de curta duração da plasticidade sináptica mediada por endocanabinoides, nas quais a despolarização pós-sináptica suprime a transmissão inibitória (DSI) ou excitatória (DSE) através da ativação pré-sináptica do CB1.
Os termos fisiológicos clássicos captam isto: supressão da inibição induzida pela despolarização, DSI, e supressão da excitação induzida pela despolarização, DSE. Ambas são formas de plasticidade sináptica de curta duração impulsionadas pela libertação de endocannabinoids e pela ativação pré-sináptica de CB1. Também ocorrem efeitos mais duradouros, incluindo depressão de longa duração mediada por endocannabinoids em algumas sinapses. Esses fenómenos são importantes porque ligam a bioquímica do recetor ao comportamento: processamento da dor, extinção do medo, aprendizagem de hábitos, apetite, controlo motor e limiar convulsivo dependem todos deste ajuste da probabilidade de libertação.
Os pormenores não são triviais. Um agonista parcial como Δ9-THC pode não mimetizar o padrão completo produzido por um breve aumento endógeno de 2-AG. Tampouco um agonista sintético total preservará necessariamente a temporização fisiológica. A dose importa. A reserva de recetor também. Numa sinapse com expressão densa de CB1, mesmo um agonista parcial pode produzir um efeito grande na libertação de neurotransmissor. Noutro tecido, o mesmo ligando pode parecer fraco.
CB2 tem uma fisiologia sináptica direta menos estabelecida do que CB1, mas também reduz cAMP e pode regular a sinalização de cálcio, vias de quinases e a libertação de mediadores inflamatórios em células imunitárias e gliais. Isso torna CB2 relevante para a comunicação neurónio-glia, especialmente em estados de doença em que a expressão do recetor se altera. O artigo recente de análise de redes indexado no PubMed em 2025/2026 tratou CB1 e CB2 como nós influentes em sinalização endocannabinoid e metabólica mais ampla, o que é um enquadramento melhor do que tratá-los como interruptores isolados.
Dessensibilização, internalização e regulação dos recetores
Como os recetores se adaptam à exposição repetida a agonistas
- Fosforilação Regiões intracelulares do recetor são modificadas por cinases do GPCR e outras cinases.
- Recrutamento de beta-arrestina As arrestinas desligam os recetores das proteínas G e podem iniciar sinalização adicional.
- Dessensibilização O recetor torna-se menos responsivo.
- Internalização Os recetores são internalizados em vias endocíticas.
- Destino após a captação Os recetores podem reciclar-se de volta para a superfície ou ser degradados.
Nenhum recetor pode ser ativado continuamente sem consequências. Para CB1 e CB2, a exposição prolongada ou repetida ao agonista conduz frequentemente à fosforilação das regiões intracelulares do recetor por GPCR kinases e outras cinases, ao recrutamento de β-arrestins, ao desacoplamento das proteínas G e, depois, à internalização através de vias endocíticas. A dessensibilização ocorre primeiro. A endocitose segue-se frequentemente. A reciclagem ou degradação vêm depois disso.
Para CB1, este ciclo regulatório é uma das principais razões pelas quais os efeitos agudos e crónicos diferem. Agonistas potentes podem desencadear rápida dessensibilização em sistemas celulares e tolerância mensurável in vivo. A regulação específica por região é importante aqui. Os recetores CB1 não dessensibilizam da mesma forma em todas as populações neuronais, o que ajuda a explicar por que a tolerância se desenvolve de maneira desigual entre os efeitos dos cannabinoids. As respostas analgésicas, a hipotermia, a perturbação da memória e os efeitos motores podem alterar-se a ritmos diferentes porque o recetor é regulado de forma diferente nos vários circuitos.
As β-arrestins não são apenas interruptores de desligar. Também podem servir de plataforma para as suas próprias cascatas de sinalização, incluindo vias da MAP kinase, razão pela qual o recrutamento de arrestina se tornou central para o agonismo enviesado. Um ligando que inibe fortemente o cAMP, mas recruta fracamente β-arrestina, pode comportar-se de forma diferente de outro que faz ambas as coisas com eficiência. Isso já não é um pormenor teórico; é uma estratégia de desenho de fármacos. A discussão de 2025 no American Journal of Psychiatry sobre o enviesamento de CB1 na esquizofrenia reflete uma lição mais ampla dos GPCR: evitar certos ramos de sinalização pode reduzir algumas limitações, mas a seletividade para uma via não garante sucesso clínico.
A própria internalização também depende do ligando. Alguns agonistas induzem endocitose extensa do recetor; outros produzem internalização limitada apesar de ativarem a proteína G. Os moduladores alostéricos complicam ainda mais o quadro ao alterar a forma como ligandos ortostéricos estabilizam os estados do recetor. É aqui que a farmacologia estrutural se cruza com a terapêutica. A revisão estrutural de 2026 deixou claro que a conformação do recetor controla em conjunto a eficácia da sinalização e a regulação do recetor, e não como tópicos separados.
Essa é a principal lição de sinalização a reter. CB1 e CB2 não são detetores simples de ligar/desligar para cannabinoids. São centros regulados cujo output muda ao longo de milissegundos a dias. Qualquer tentativa séria de os atingir, seja para epilepsia, dor, psicose ou doença inflamatória, tem de considerar o acoplamento Gi/o, os segundos mensageiros, o controlo de canais iónicos, a localização sináptica e o facto de o recetor se adaptar a ser estimulado.
Sinalização enviesada: porque é que um recetor pode produzir diferentes resultados biológicos
A antiga visão da farmacologia dos cannabinoid tratava um recetor como um interruptor de luz: os agonistas ligam-no, os antagonistas desligam-no, e tudo o resto decorre de onde esse recetor estiver expresso. Essa visão não é adequada para CB1 ou CB2. Não explica porque é que dois ligandos que atuam no mesmo recetor podem produzir efeitos comportamentais, cognitivos, inflamatórios ou terapêuticos muito diferentes. Também não explica porque é que a descoberta de fármacos dirigidos aos recetores cannabinoid tem produzido repetidamente compostos que pareciam promissores in vitro, mas se revelaram dececionantes, intoleráveis ou clinicamente ambíguos.
Isso tem importância muito para além da teoria académica dos recetores. A World Health Organization estimou que 200 milhões de pessoas usaram cannabis em 2019, cerca de 4% da população mundial com idades entre os 15 e os 64 anos. A esquizofrenia afeta cerca de 24 milhões de pessoas em todo o mundo. Perante esse enquadramento, a farmacologia de CB1 não é uma questão periférica. Situa-se na interseção entre saúde pública, psiquiatria e desenho de fármacos. A U.S. FDA, até 2025, tinha aprovado um produto medicinal derivado de cannabis e três produtos relacionados com cannabis, um número reduzido face à escala do interesse clínico. Uma razão para o progresso ter sido mais lento do que a discussão pública sugere é que a sinalização dos recetores cannabinoid não se resume à ocupação simples do recetor. Trata-se de seleção de vias.
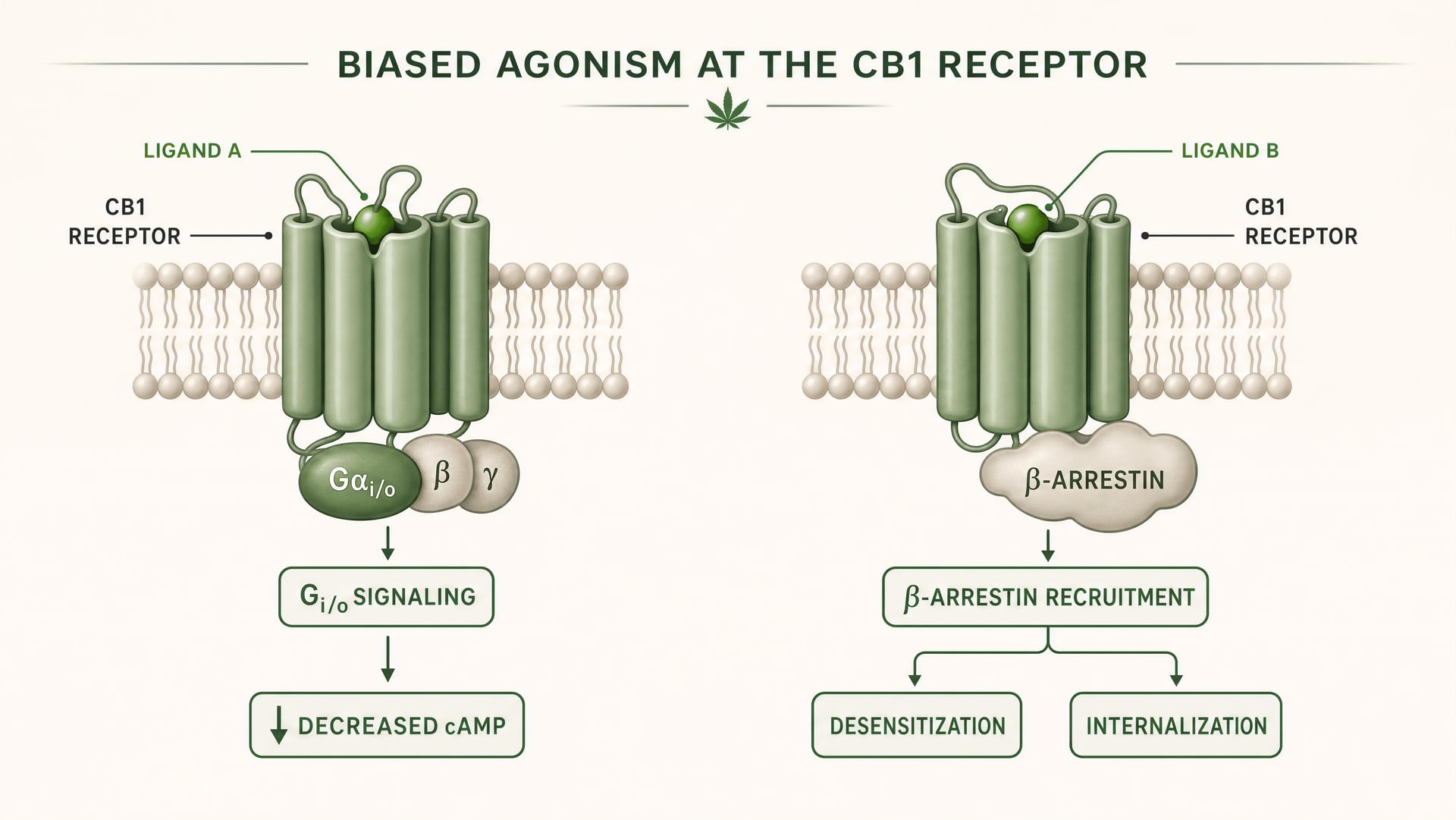
O que significa agonismo enviesado na farmacologia dos GPCR
CB1 e CB2 são recetores acoplados à proteína G da classe A. O trabalho fundador de Allyn Howlett estabeleceu CB1 como um recetor cannabinoid acoplado a Gi/o, ajudando a mover o campo de uma farmacologia vaga para mecanismos definidos por recetor. Mas o acoplamento a Gi/o é apenas o início da história. Depois de um ligando se ligar, o recetor pode adotar mais do que uma forma ativa, e essas formas não sinalizam de maneira idêntica. Algumas conformações do recetor favorecem a ativação da proteína G. Outras recrutam com maior intensidade as beta-arrestinas. Alguns estados promovem fosforilação do recetor, dessensibilização ou internalização. Outros produzem sinalização mais duradoura a partir da membrana plasmática ou de compartimentos endossómicos.
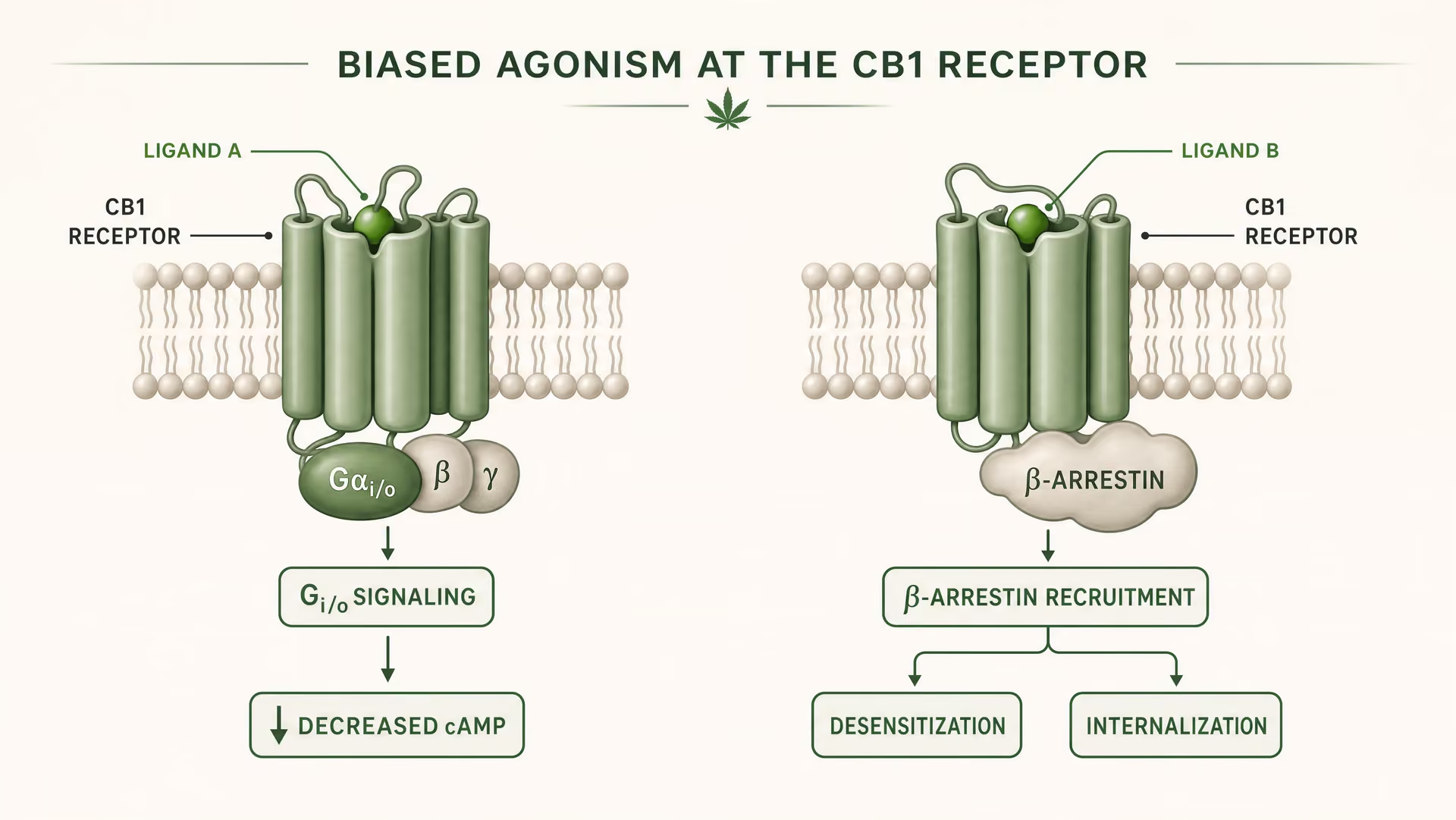{kind=link}
Agonismo enviesado Uma propriedade de um ligando que favorece algumas vias de sinalização a jusante em detrimento de outras no mesmo recetor.
Em termos simples, isto é agonismo enviesado: ligandos diferentes estabilizam conformações diferentes do recetor, e essas conformações favorecem vias a jusante distintas. Um recetor não está apenas ligado ou desligado. Está conformacionalmente orientado.
Para CB1, isto é especialmente importante porque o recetor se encontra num ambiente de sinalização denso, plástico e altamente dependente do tipo celular. Num terminal glutamatérgico cortical, um ligando pode reduzir a libertação do neurotransmissor através da inibição da adenilato ciclase mediada por Gi/o e da modulação de canais iónicos. Num interneurónio GABAérgico, o mesmo recetor pode deslocar o equilíbrio local do circuito numa direção muito diferente. Se o ligando também promover forte recrutamento de beta-arrestina, o recetor pode internalizar mais rapidamente, encurtando um efeito enquanto abre outro. O tempo muda. A localização do sinal muda. A resposta fisiológica muda.
Isto não é um preciosismo teórico. A revisão estrutural de 2026 em Frontiers in Chemical Biology sobre recetores cannabinoid deixa o ponto claro: a seletividade dos ligandos em CB1 e CB2 depende de diferenças estruturais ao nível do recetor que alteram a ligação, a eficácia de sinalização e a regulação do recetor. A palavra-chave aí é regulação. Um ligando pode ter afinidade semelhante, mas diferir em eficácia, recrutamento de arrestina, tempo de permanência ou propensão para desencadear dessensibilização. O estudo indexado no PubMed de 2025/2026 sobre o mecanismo dinâmico da seletividade entre subtipos vai na mesma direção ao argumentar que a seletividade emerge de dinâmicas conformacionais, e não de um modelo estático de chave-fechadura. Os endocannabinoid, os phytocannabinoids e os ligandos sintéticos não devem, portanto, ser agrupados indiscriminadamente. Anandamide, descoberta por Raphael Mechoulam e Lumír Hanuš, não se comporta como delta-9-tetrahydrocannabinol, e nenhum deles se comporta como uma sonda sintética altamente otimizada.
A sinalização enviesada também explica porque é que os moduladores alostéricos atraem tanto interesse. Um ligando alostérico pode não ativar diretamente CB1 da mesma forma que um agonista ortostérico, mas pode remodelar as preferências de sinalização do recetor, amplificando uma via e atenuando outra. Isso abre uma via para um controlo fino. Em princípio.
Sinalização enviesada de CB1 como direção de investigação na esquizofrenia
O artigo de 2025 no American Journal of Psychiatry apresenta o argumento mais forte até à data de que a sinalização enviesada de CB1 não é apenas um conceito de farmacologia, mas uma estratégia terapêutica plausível para a esquizofrenia. Esse argumento merece atenção porque a investigação em esquizofrenia tem abordado os cannabinoid sobretudo através da epidemiologia, da associação de risco ou de advertências amplas sobre psicose. O artigo do AJP muda o enquadramento. Pergunta se o problema não será “cannabinoid” em geral, ou mesmo a “ativação de CB1” em geral, mas sim quais estados de sinalização de CB1 estão a ser ativados, em que circuitos e durante quanto tempo.
Essa é uma pergunta muito melhor.
CB1 é um dos GPCR mais abundantes no cérebro, com expressão elevada no córtex, hipocampo, gânglios da base e cerebelo, mas a abundância por si só não explica os efeitos clínicos. A esquizofrenia envolve desregulação da saliência, da cognição, da perceção e da coordenação em rede em sistemas corticais e subcorticais. Um recetor posicionado para modular a atividade dos circuitos glutamatérgicos, GABAérgicos e relacionados com a dopamina é, portanto, relevante por definição. O artigo do AJP defende que ligandos enviesados de CB1 poderão separar efeitos de circuito terapeuticamente úteis de limitações como compromisso cognitivo, ansiedade, disforia ou respostas psicotomiméticas.
Trata-se de uma afirmação ambiciosa, mas não é especulação vazia. Segue a lógica do campo mais vasto dos GPCR, em que o enviesamento de vias já mudou a forma como os investigadores pensam sobre fármacos dos recetores opioides, da angiotensina e da dopamina. A esperança translacional em CB1 é que determinados resultados de sinalização possam melhorar a função das redes corticais ou atenuar estados de circuito aberrantes sem reproduzir o perfil completo de efeitos adversos associado ao agonismo de alta eficácia de CB1.
A esquizofrenia é um bom caso de teste porque a fasquia clínica é elevada. Um candidato a fármaco não pode apenas alterar o comportamento num ensaio em roedor. Tem de evitar agravar psicose, sedação e perturbação cognitiva em pessoas já vulneráveis a esses problemas. Isso torna a seletividade de via mais do que uma preferência de química medicinal. Torna-se um requisito de segurança.
O enquadramento do AJP também ajuda a corrigir uma simplificação frequente nas discussões sobre cannabis. Delta-9-THC é um phytocannabinoid com atividade agonista parcial em CB1, mas os seus efeitos refletem dose, timing, reserva de recetor, tónus endocannabinoid local e envolvimento de vias em diferentes populações neuronais. Um ligando sintético de CB1 concebido para favorecer uma via intracelular específica pode parecer muito diferente de THC, mesmo que ambos “atinjam CB1”. O inverso também é verdadeiro: dois compostos que melhorem um desfecho pré-clínico relevante para a esquizofrenia podem divergir de forma acentuada em cognição ou afeto se um deles conduzir sinalização fortemente associada a arrestina e o outro não. A identidade do recetor, por si só, não consegue prever todo o fenótipo.
Porque é que a seletividade de via é importante para a segurança e a eficácia
A seletividade de via é importante porque a eficácia não é uma dimensão única. Um fármaco cannabinoid pode ser potente e, ainda assim, clinicamente fraco. Pode ser seletivo para CB1 e, mesmo assim, falhar. Pode evitar completamente CB2 e, ainda assim, produzir efeitos imunitários ou metabólicos indesejáveis através de interações em rede. A análise de rede integrativa de 2025/2026 indexada no PubMed identificou CB1 e CB2 como nós altamente influentes no endocannabinoid system e mapeou a sua sinalização para vias metabólicas. Essa perspetiva sistémica é essencial. Os recetores não operam isoladamente, e o enviesamento de via num nó pode repercutir-se em programas fisiológicos mais amplos.
Para CB1, as preocupações de segurança são óbvias. Uma forte ativação central de CB1 pode provocar défice de memória, alteração da perceção, ansiedade, taquicardia e, em indivíduos suscetíveis, efeitos relacionados com psicose. Qualquer programa terapêutico dirigido a dor, apetite, humor, dependência ou esquizofrenia tem de enfrentar esse perfil de risco. Um ligando que preserve um efeito sináptico desejado mediado por Gi/o enquanto limita a dessensibilização mediada por beta-arrestina ou outras cascatas de sinalização adversas poderá, em teoria, alargar a janela terapêutica. Mas “em teoria” é importante. Muitos programas de ligandos enviesados na farmacologia dos GPCR demonstraram que o enviesamento medido num sistema de ensaio nem sempre prevê resultados in vivo. O fundo celular, a densidade do recetor, a expressão de efetores e a cinética podem alterar o enviesamento aparente.
CB2 oferece um paralelo de prudência. A revisão de 2026 em Frontiers in Behavioral Neuroscience descreve uma atualização dos últimos 3 anos na qual a sinalização de CB2 ganhou atenção em perturbações do sistema nervoso central através de ligações a mecanismos neuroinflamatórios e neurodegenerativos. Isso contraria diretamente a velha ideia de que CB2 é irrelevante para o cérebro. Ainda assim, visar CB2 não garante, por si só, um medicamento anti-inflamatório útil. A distribuição é mais gradual do que a antiga divisão cérebro-versus-corpo, e as consequências da sinalização continuam a depender do ligando e do contexto.
A lição prática é, portanto, direta: a seletividade do subtipo de recetor é necessária, mas não suficiente. A seletividade de via pode ser a diferença entre um cannabinoid que parece terapêutico, um que é intoxicante e um que falha nos ensaios porque não consegue separar benefício de efeito adverso. Para CB1, especialmente em psiquiatria, é provável que essa distinção determine se o recetor permanece um aviso ou se se torna um alvo farmacológico viável.
Biologia estrutural de CB1 e CB2: como a forma determina a seletividade
A biologia estrutural mudou a forma como os receptores de cannabinoid são discutidos. A antiga simplificação — CB1 explica a intoxicação, CB2 explica a inflamação — ignora o facto de ambos os receptores serem receptores acoplados à proteína G de classe A, cujo comportamento depende da forma, do movimento, da profundidade de ligação e dos parceiros de sinalização disponíveis numa determinada célula. Isso tem implicações muito para além da farmacologia básica. Cannabis foi usada por um número estimado de 200 milhões de pessoas em 2019, ou 4% da população global com idades entre os 15 e os 64 anos, segundo a WHO, mas a FDA ainda lista apenas um medicamento derivado de cannabis e três medicamentos relacionados com cannabis como aprovados em 2025. Uma das razões para essa diferença é estrutural: é difícil criar ligandos de cannabinoid que atinjam o recetor certo, da forma certa, durante o período certo.
A revisão da Frontiers in Chemical Biology de 2026 torna este ponto claro. CB1 e CB2 não diferem apenas no local onde são expressos. Diferem na arquitetura das suas cavidades de ligação ao ligando, na forma e flexibilidade dos seus loops extracelulares, no empacotamento das suas hélices transmembranares e nos estados conformacionais que preferem após a ligação de um ligando. Essas características influenciam não apenas a seletividade, mas também a eficácia, a dessensibilização, a internalização e o enviesamento de via.
O que os estudos estruturais revelam sobre as cavidades do recetor
Uma cavidade ortostérica é a principal cavidade de ligação onde ligandos endógenos como anandamide e 2-arachidonoylglycerol, fitocannabinoids como THC e muitos ligandos sintéticos estabelecem o seu contacto primário. Em CB1 e CB2, essa cavidade situa-se no interior do feixe de sete hélices transmembranares, parcialmente coberto por loops extracelulares que podem abrir o acesso ou restringi-lo.
Estruturas de cryo-EM e de raio X obtidas ao longo dos últimos anos mostraram que os receptores de cannabinoid não se comportam como fechaduras rígidas à espera de uma chave. São melhor entendidos como alvos móveis com formas preferenciais. A revisão da Frontiers in Chemical Biology de 2026 sublinha que as cavidades ortostéricas de CB1 e CB2 são suficientemente semelhantes para ligar classes sobrepostas de ligandos, mas suficientemente diferentes em tamanho, identidade dos resíduos e flexibilidade local para alterar a afinidade e o resultado da sinalização. É por isso que compostos estreitamente relacionados podem separar-se farmacologicamente. Uma pequena alteração na dimensão de um substituinte, na polaridade ou no comprimento da cadeia pode modificar a profundidade com que um ligando penetra na cavidade, as hélices que empurra e se o recetor adota um estado favorável à proteína G ou favorável à arrestina.
CB1 tem sido especialmente informativo do ponto de vista estrutural porque existem agora muitos modelos de alta resolução em estados inativo e ativo. Um tema recorrente é que a sua cavidade é expansiva e hidrofóbica, acomodando a natureza lipofílica de muitos cannabinoids. O loop extracelular 2 e as porções superiores de várias hélices ajudam a moldar a entrada. As hélices transmembranares são os sete segmentos que atravessam a membrana e que constituem o núcleo do recetor; quando um ligando se liga, estas hélices podem deslocar-se relativamente umas às outras. O movimento mais importante do ponto de vista farmacológico ocorre geralmente no lado intracelular, onde o movimento para fora da hélice 6 ajuda a criar um local de ancoragem para as proteínas Gi/o. Esse deslocamento é uma das marcas da ativação do recetor.
CB2 partilha o mesmo dobramento global de GPCR, mas a revisão da Frontiers argumenta que diferenças específicas de aminoácidos em torno da cavidade e das regiões de loop fornecem aos químicos medicinais pontos de intervenção exploráveis para seletividade. A questão não é que uma cavidade seja simplesmente “semelhante ao cérebro” e a outra “semelhante ao sistema imunitário”. A questão é geométrica e energética. Resíduos distintos alteram o contorno da cavidade, as opções locais de ligação por hidrogénio, o empilhamento aromático e a flexibilidade dos canais de acesso através dos quais os ligandos entram a partir da membrana.
Um estudo indexado no PubMed de 2025/2026 sobre o mecanismo dinâmico da seletividade por subtipo avançou este raciocínio ao defender que a seletividade de endocannabinoid não é apenas uma questão de afinidade de ligação estática. A dinâmica conformacional importa. Em termos simples, um recetor pode amostrar várias formas antes e depois da ligação do ligando, e alguns ligandos estabilizam melhor uma forma seletiva do que outros. Isso ajuda a explicar por que lípidos endógenos, fitocannabinoids e ligandos sintéticos podem mostrar preferências diferentes por subtipo, mesmo quando as suas estruturas parecem relacionadas no papel.
Determinantes da seletividade de ligandos entre CB1 e CB2
A seletividade começa na química de contacto, mas não termina aí. A revisão da Frontiers in Chemical Biology enquadra a seletividade como o produto de diferenças estruturais ao nível do recetor que afetam simultaneamente a ligação, a eficácia da sinalização e a regulação. Esse é o enquadramento correto. Um ligando pode ser seletivo para CB2 num ensaio de ligação com radioligando e, ainda assim, perder a sua vantagem prática se também induzir estados do recetor que causem tolerância rápida ou sinalização fraca em células relevantes para a doença.
Várias características estruturais surgem repetidamente. Primeiro, a composição de aminoácidos da cavidade ortostérica difere o suficiente entre CB1 e CB2 para alterar a forma como o grupo polar da molécula, o núcleo e a cadeia hidrofóbica são acomodados. Segundo, os loops extracelulares ajudam a moldar a entrada e a orientação. Terceiro, as regiões superiores e médias das hélices transmembranares podem enviesar o recetor para conjuntos de estados ativos ligeiramente diferentes. Um estado conformacional é simplesmente uma das possíveis formas do recetor num dado momento. Diferentes ligandos não se limitam a ligar-se a um recetor; estabilizam um subconjunto dessas formas.
É por isso que a seletividade por subtipo é frequentemente modesta para cannabinoids naturais. THC, por exemplo, interage com ambos os recetores. Anandamide e 2-AG também atuam sobre ambos, embora com diferenças dependentes do contexto em potência, eficácia e metabolismo. Os ligandos sintéticos têm sido mais úteis para dissecar relações entre estrutura e seletividade porque os químicos podem alterar sistematicamente características como o comprimento da cadeia lateral, restrições do anel e substituintes polares. Mesmo assim, a separação limpa é difícil. CB1 e CB2 são suficientemente homólogos para que um composto concebido para um deles muitas vezes mantenha atividade relevante no outro.
Isto tem consequências práticas. Há muito que os investigadores procuram agonistas seletivos de CB2 na esperança de obter efeitos anti-inflamatórios ou analgésicos sem efeitos adversos centrais fortes mediados por CB1. Por vezes, essa estratégia funciona farmacologicamente, mas não é um passe livre. A revisão da Frontiers in Behavioral Neuroscience de 2026 sublinha que CB2 tem atraído atenção em doenças do sistema nervoso central ao longo dos “últimos 3 anos”, contrariando a visão simplista de que CB2 é irrelevante para o cérebro. Assim, mesmo um ligando “periférico” de CB2 não pode ser interpretado com base em mapas do recetor desatualizados.
Porque é que eficácia e regulação também são questões estruturais
Potência responde à pergunta de quanta quantidade de ligando é necessária. Eficácia pergunta o que é que esse ligando faz ao recetor depois de se ligar. A biologia estrutural liga os dois conceitos, mas não os confunde. Dois ligandos podem ocupar a mesma cavidade e produzir respostas muito diferentes porque estabilizam conformações ativas distintas.
Para CB1, isto é central para o pensamento terapêutico atual. O artigo do American Journal of Psychiatry de 2025 argumenta que a sinalização enviesada em CB1 pode ser uma estratégia plausível para a esquizofrenia, uma perturbação que afeta cerca de 24 milhões de pessoas em todo o mundo. Agonismo enviesado significa que um ligando favorece uma via downstream em detrimento de outra, muitas vezes a sinalização Gi/o em vez do recrutamento de beta-arrestin, ou vice-versa. Estruturalmente, esse enviesamento resulta do facto de o ligando deslocar a face intracelular do recetor para uma forma que se adapta melhor a um parceiro de sinalização do que a outro. Isso não é uma ideia abstrata. É um alvo de química medicinal.
As beta-arrestins são importantes porque contribuem para a dessensibilização e a internalização. Dessensibilização significa que o recetor se torna menos responsivo após ativação repetida ou prolongada. Internalização significa que o recetor é puxado para o interior da célula a partir da membrana de superfície. Ambos os processos são influenciados pela conformação do recetor. Alguns ligandos ativam fortemente as proteínas G mas recrutam arrestins apenas de forma fraca; outros fazem ambos. Um medicamento de CB1 tolerável do ponto de vista comportamental pode precisar precisamente deste tipo de separação, em vez de simples bloqueio ou agonismo total.
A modulação alostérica acrescenta outra camada. Um ligando alostérico liga-se fora da cavidade ortostérica e altera a forma como o recetor responde a outros ligandos. Estruturalmente, isto é apelativo porque pode permitir um controlo mais fino da forma do recetor do que bombardear a cavidade principal com um agonista de elevada eficácia. Para a farmacologia de cannabinoid, isso pode significar preservar a sinalização útil enquanto se reduzem os efeitos adversos associados a uma ativação excessiva de CB1.
A lição mais ampla é direta. A forma do recetor não é um detalhe secundário. É a explicação para o facto de fitocannabinoids, endocannabinoids e compostos sintéticos poderem produzir perfis clínicos diferentes, mesmo quando visam a mesma família de recetores. Como CB1 e CB2 ocupam nós influentes nas análises de rede do sistema endocannabinoid, como reportou um estudo de sistemas indexado no PubMed de 2025/2026, erros de seletividade ou eficácia podem propagar-se por muitas vias. A informação estrutural não garante um fármaco bem-sucedido. Mas explica por que razão a seletividade, a potência e a tolerabilidade continuam tão difíceis de harmonizar.
Endocanabinóides e selectividade de subtipo: anandamida, 2-AG e comportamento dinâmico dos receptores
A farmacologia dos endocanabinóides é frequentemente explicada com uma simplificação que parece elegante, mas é enganadora: a anandamida e o 2-arachidonoylglycerol (2-AG) são tratados como se fossem os “agonistas padrão” integrados do organismo, úteis sobretudo como pontos de referência para o que CB1 e CB2 fazem normalmente. Esse enquadramento omite o facto central. Estes não são hormonas estáveis, livremente circulantes, que encontrem receptores em condições uniformes. São mediadores lipídicos de curta duração, produzidos sob demanda, nas membranas, por sistemas enzimáticos específicos de cada célula, sendo depois rapidamente degradados. A sua preferência aparente por determinados receptores pode variar com a composição da membrana, as conformações dos receptores, a atividade enzimática local e o momento da libertação.
Isso importa para além da teoria académica dos receptores. Estima-se que o cannabis foi utilizado por 200 milhões de pessoas em 2019, ou 4% da população mundial com idades entre os 15 e os 64 anos, segundo a WHO, e os medicamentos direcionados para cannabinoid já estão na clínica: a FDA afirmou em 2025 que aprovou um produto medicamentoso derivado de cannabis e três produtos medicinais relacionados com cannabis. Se os ligandos endógenos não se comportam como simples controlos de manual, então comparar fitocanabinoides ou candidatos a fármacos com eles não é uma tarefa linear.
Porque os ligandos endógenos não são compostos de referência simples
A anandamida, identificada em 1992 por Devane, Hanus, Breuer, Mechoulam e colegas, e o 2-AG, estabelecido alguns anos mais tarde como outro endocanabinóide importante, diferem em mais do que tabelas de potência. Resultam de vias biossintéticas diferentes, atingem os receptores em escalas temporais distintas e são eliminados por enzimas diferentes. A anandamida é gerada sobretudo a partir de N-arachidonoyl phosphatidylethanolamine através de vias envolvendo NAPE-PLD, embora existam vias alternativas. O 2-AG é produzido em grande parte a partir de diacilglicerol por DAGLα e DAGLβ. Estes não são pormenores bioquímicos menores; determinam onde e quando ocorre a ativação do receptor.
Até a afirmação mais antiga de que a anandamida é um ligando com preferência para CB1, enquanto o 2-AG é um agonista total em CB1 e CB2, pode tornar-se demasiado simplista quando transportada de sistemas purificados para tecido vivo. A anandamida é também um substrato para fatty acid amide hydrolase, ou FAAH, e pode interagir com alvos não cannabinoid como TRPV1 em determinadas condições. O 2-AG está presente no tecido cerebral em níveis basais muito superiores aos da anandamida em muitas medições e é hidrolisado sobretudo por monoacylglycerol lipase, ou MAGL, contribuindo também ABHD6 e ABHD12. Assim, a “entrada” para o receptor que cada ligando fornece não depende apenas da afinidade ou da eficácia em CB1 versus CB2. Depende de o ligando ser produzido na sinapse, no interior de uma célula imunitária, próximo de uma membrana intracelular ou num compartimento rico em enzimas degradativas.
O trabalho de farmacologia de receptores de Allyn Howlett ajudou a estabelecer CB1 como um GPCR autêntico, acoplado em grande medida às proteínas Gi/o. Esse enquadramento continua essencial, mas já não suporta um modelo simples de fármaco de referência endógeno. CB1 e CB2 podem sinalizar através da inibição da adenylyl cyclase, da regulação de canais iónicos, de vias MAP kinase, do recrutamento de beta-arrestin, da internalização do receptor e da dessensibilização. Um ligando que parece “mais fraco” num ensaio pode produzir sinalização mais prolongada num tecido onde a degradação é lenta, ou pode favorecer uma via a jusante em detrimento de outra. O artigo de 2025 do American Journal of Psychiatry sobre sinalização enviesada de CB1 sublinha este ponto num contexto de doença: a questão terapêutica em perturbações como a esquizofrenia, que afeta cerca de 24 milhões de pessoas em todo o mundo, não é apenas se CB1 é ativado, mas como.
Mecanismos dinâmicos da selectividade de subtipo
O recente estudo sobre selectividade de subtipo indexado no PubMed (PMID: 41962866) é importante porque afasta o campo de uma narrativa de chave-fechadura. A sua mensagem central é que a selectividade dos endocanabinóides pode emergir de mecanismos dinâmicos. Por outras palavras, a preferência por um subtipo de receptor não é explicada apenas por uma diferença fixa na força com que um ligando se liga a CB1 versus CB2. O complexo ligando-receptor amostra múltiplos estados conformacionais, e esses estados diferem entre CB1 e CB2.
Essa conclusão é coerente com a revisão de 2026 em Frontiers in Chemical Biology, que defende que as diferenças estruturais entre CB1 e CB2 moldam a ligação, a eficácia de sinalização e a regulação do receptor. Os receptores partilham uma semelhança de sequência substancial, mas pequenas diferenças na arquitetura da bolsa ortostérica, nos loops extracelulares, nos movimentos das hélices transmembranares e nas superfícies de acoplamento intracelular podem enviesar o que acontece após a ligação do ligando. Um ligando pode entrar em bolsas semelhantes nos dois receptores, mas estabilizar um estado mais competente para sinalização num subtipo, ou favorecer diferentes transições entre estados inativo, intermédio e ativo.
Para os endocanabinóides, isto é especialmente plausível porque são lípidos flexíveis, e não moldes sintéticos rígidos. A flexibilidade é uma variável farmacológica. A anandamida pode adotar múltiplas conformações, e o 2-AG é igualmente dinâmico nas membranas. A questão deixa então de ser apenas “Liga-se ou não?” e passa a ser “Quais os estados do receptor que favorece, durante quanto tempo esses estados são ocupados, e que proteínas estão disponíveis para o acoplamento seguinte?” Um neurónio rico em CB1 e uma célula microglial que exprime CB2 não apresentam o mesmo ambiente de sinalização, mesmo antes de se considerarem diferenças na densidade de receptores.
Esta visão dinâmica também enfraquece a antiga suposição de que os ligandos endógenos definem uma única linha de base fisiológica contra a qual THC, cannabidiol ou cannabinoid sintéticos possam ser classificados. THC é um fitocanabinoide com o seu próprio padrão de eficácia e comportamento cinético. Os endocanabinóides são sinais desencadeados por eventos, muitas vezes fásicos e locais. Um agonista sintético de CB2 pode parecer seletivo num ensaio recombinante, mas produzir resultados inesperados em tecido se a reserva de receptores, o colesterol da membrana, a formação de heterómeros ou o processamento por beta-arrestin forem diferentes. A seletividade no papel não é determinante.
Como a síntese e a degradação locais moldam o engajamento do receptor
In vivo, a sinalização dos endocanabinóides é governada tanto pela enzimologia como pela farmacologia dos receptores. Em muitas sinapses centrais, o 2-AG é produzido pós-sinapticamente em resposta a aumentos de cálcio ou à ativação de receptores acoplados a Gq/11, deslocando-se depois retrogradamente para os receptores CB1 pré-sinápticos e suprimindo a libertação de neurotransmissores. Este é um mecanismo temporal medido em segundos, não um tom de fundo difuso. O sinal termina quando MAGL e hidrolases relacionadas removem o 2-AG. A anandamida comporta-se frequentemente de forma diferente, com menor abundância tecidular, condições de libertação distintas e maior sensibilidade à degradação controlada por FAAH.
Isto significa que alterar uma enzima pode modificar o engajamento do receptor sem alterar o próprio receptor. A inibição de FAAH aumenta o tónus da anandamida, mas o resultado fisiológico depende de onde a anandamida é produzida, de se TRPV1 também está presente e de se os receptores CB1 nesse microdomínio estão dessensibilizados ou internalizados. A inibição de MAGL pode elevar de forma marcada o 2-AG, mas a elevação crónica pode induzir dessensibilização de CB1 em alguns sistemas. Mais ligando nem sempre significa sinalização mais útil.
O estudo de sistemas biológicos indexado como PMID: 42129940 reforça esta visão mais ampla ao colocar CB1 e CB2 entre nós influentes ligados a vias metabólicas, e não como interruptores isolados. Essa é a escala correta para compreender a ação dos endocanabinóides. A disponibilidade do ligando, o acesso à membrana, a conformação do receptor, o acoplamento à proteína G, o recrutamento de beta-arrestin e a degradação interagem todos entre si. O mesmo acontece com os gradientes tecidulares. CB2 não está simplesmente “fora do cérebro”, e CB1 não é apenas o “receptor da psicoatividade”. A sinalização dos endocanabinóides é local, condicional e cinética. Qualquer descrição da selectividade de subtipo que ignore essas características irá interpretar mal tanto a fisiologia como o desenvolvimento de medicamentos.
O sistema endocanabinoide como uma rede, não um diagrama de dois receptores
O sistema endocanabinoide faz pouco sentido se for representado como dois círculos rotulados CB1 e CB2 com setas a apontar para o exterior. Esse esquema ajudou a área nos seus primórdios, sobretudo após o trabalho de farmacologia de recetores de Allyn Howlett e a identificação da anandamida por Raphael Mechoulam e Lumír Hanuš, mas hoje oculta mais do que revela. Os riscos práticos são elevados. A World Health Organization estimou que 200 milhões de pessoas usaram cannabis em 2019, cerca de 4% da população mundial com idades entre os 15 e os 64 anos, enquanto a U.S. FDA afirma que, em 2025, estão aprovados um produto farmacêutico derivado de cannabis e três produtos farmacêuticos relacionados com cannabis. Um sistema de recetores que toca em tantas exposições e em tantas alegações terapêuticas não pode ser tratado como um simples par de recetores, um cerebral e outro imunitário.
Uma perspetiva sistémica começa por um facto básico: os endocanabinoides são lípidos produzidos sob demanda, degradados rapidamente e inseridos no mesmo espaço bioquímico que os mediadores inflamatórios, as vias de remodelação de membranas e os circuitos de feedback sináptico. A anandamida e o 2-arachidonoylglicerol não são “mensagens” isoladas. São produzidos a partir de precursores da membrana, competem com outras vias lipídicas por substratos e atuam em tecidos onde a densidade dos recetores, o tipo celular, a expressão enzimática e os parceiros de sinalização diferem. É por isso que o mesmo ligando pode parecer ansiolítico num contexto, intoxicante noutro, anti-inflamatório noutro ainda, e clinicamente pouco convincente num ensaio.
O que a análise integrativa de redes acrescenta
O estudo de análise integrativa de redes indexado no PubMed, publicado em 2025/2026 (PMID: 42129940), é útil porque desloca a atenção de alvos únicos para a organização do sistema. Em vez de perguntar apenas que ligando se liga a CB1 ou CB2, os autores mapeiam o sistema endocanabinoide como uma rede de interações que inclui recetores, enzimas biossintéticas e degradativas, intermediários lipídicos e ligações a nível de via com inflamação, metabolismo e sinalização neuronal. Nesse enquadramento, a influência não é definida apenas pela abundância do recetor. É definida pela conectividade e pela forma como as perturbações se propagam pela rede.
Isso importa para a interpretação. Se um fármaco altera a sinalização de CB1, o efeito a jusante não se limita à inibição mediada por Gi/o da adenilato ciclase. Pode alterar a libertação de neurotransmissores, modificar o comportamento dos canais de cálcio e potássio, alterar o recrutamento de beta-arrestina, desencadear a internalização do recetor e, indiretamente, modificar o fluxo de substratos lipídicos que também alimentam eicosanoides e outras vias inflamatórias. Um modelo de rede capta melhor estas consequências entre sistemas do que um gráfico de ocupação de recetores.
É também aqui que o velho resumo “CB1 no cérebro, CB2 nas células imunitárias” deixa de funcionar. A distribuição é graduada, específica do tipo celular e dependente do estado. CB1 é abundante em muitas populações neuronais, mas não de forma uniforme em todo o cérebro, e aparece também em tecidos periféricos relevantes para o equilíbrio energético, a nociceção e a função dos órgãos. CB2 está enriquecido em linhagens imunitárias, mas trabalhos mais recentes no sistema nervoso central afastaram-se claramente da ideia de que é irrelevante para a biologia cerebral. Uma revisão de 2026 na Frontiers in Behavioral Neuroscience descreveu isto como “an update over the last 3 years”, sublinhando a sinalização de CB2 em contextos neuroinflamatórios e neurodegenerativos. O ponto não é que CB2 tenha subitamente passado a ser “também o recetor do cérebro”. O ponto é que o significado do recetor depende de quais células estão ativas, lesionadas, inflamadas ou em adaptação.
A análise de redes ajuda a explicar por que razão os fármacos dirigidos a recetores frequentemente produzem efeitos mais amplos do que o esperado. Um fitocanabinoide ativo em CB1, como o delta-9-THC, um endocanabinoide como a anandamida e um agonista sintético podem todos ativar o mesmo recetor e, ainda assim, gerar resultados temporais e espaciais diferentes. A classe do ligando importa. Importam também a eficácia, o tempo de residência, o metabolismo e o acesso a diferentes conformações do recetor.
CB1 e CB2 como nós influentes
O estudo integrativo de redes identifica CB1 e CB2 como nós altamente influentes, não porque operem isoladamente, mas porque se situam em pontos de controlo onde convergem muitas vias. Em termos de rede, nós influentes são locais onde alterações locais podem propagar-se amplamente. Em termos de farmacologia, são recetores cujo estado de ativação pode redirecionar simultaneamente a sinalização sináptica, as respostas imunitárias e os programas de stress celular.
CB1 é o exemplo mais claro de porque “influente” não significa “simples”. Sinaliza sobretudo através de proteínas Gi/o, mas isso é apenas o início. Dependendo do ligando e do contexto, CB1 pode recrutar beta-arrestinas, sofrer dessensibilização dependente de fosforilação, internalizar-se, reciclar ou permanecer persistentemente ativo na membrana. O artigo do American Journal of Psychiatry de 2025 argumenta que a sinalização enviesada em CB1 é uma estratégia terapêutica plausível para a esquizofrenia, uma perturbação que a WHO diz afetar cerca de 24 milhões de pessoas em todo o mundo. Essa proposta só faz sentido se CB1 for entendido como um eixo de sinalização com saídas separáveis, e não como um interruptor de ligação/desligação para intoxicação. Um ligando que favoreça uma via de sinalização em detrimento de outra pode preservar alguns efeitos terapêuticos enquanto reduz os adversos. Pode, não irá necessariamente. O enviesamento pode reduzir responsabilidades, mas não apaga o facto de CB1 estar inserido em circuitos que regulam cognição, recompensa, controlo motor, processamento da dor e apetite.
CB2 mostra um problema semelhante, mas a partir da direção oposta. Durante muito tempo foi apresentado como o alvo mais seguro devido à menor associação com efeitos psicoativos agudos. Esse enquadramento é demasiado otimista. Ligandos dirigidos a CB2 ainda podem produzir resultados desapontantes ou mistos se a sinalização relevante para a doença depender do estado celular, da inducibilidade do recetor, do tónus inflamatório ou da interação cruzada com enzimas metabólicas. A recente revisão estrutural na Frontiers in Chemical Biology, publicada em 2026, enfatiza que a seletividade entre “CB1 e CB2” depende de diferenças estruturais ao nível do recetor que alteram a ligação, a eficácia e a regulação do recetor. O estudo de 2025/2026 indexado no PubMed sobre seletividade de subtipos (PMID: 41962866) vai mais longe ao argumentar que a seletividade é dinâmica, moldada pelo comportamento conformacional e não por um ajuste estático do tipo fechadura-chave. Esta é uma razão forte para deixar de falar de compostos seletivos para CB1 ou seletivos para CB2 como se a seletividade fosse uma propriedade fixa com consequências biológicas fixas.
Ligações entre a sinalização dos recetores e as vias metabólicas
A contribuição mais valiosa da perspetiva de rede é que volta a ligar a farmacologia dos recetores ao metabolismo lipídico. A sinalização endocanabinoide assenta na química relacionada com o ácido araquidónico. A anandamida e o 2-AG são gerados e degradados por sistemas enzimáticos que também influenciam a disponibilidade de lípidos bioativos envolvidos na inflamação e na resposta tecidular. Um fármaco que inibe FAAH ou MAGL não “aumenta apenas os endocanabinoides”. Redistribui o tráfego de sinalização por uma rede metabólica.
Isso pode ser útil. Mas também pode correr mal. A elevação de 2-AG, por exemplo, pode potenciar a sinalização de CB1 ou CB2 em alguns compartimentos enquanto altera vias relacionadas com prostaglandinas noutros, através de desvio de substrato ou oxidação a jusante. A implicação clínica é clara: estratégias dirigidas a recetores e estratégias dirigidas a enzimas podem ter consequências em todo o sistema, mesmo quando o objetivo do desenho é estreito. Esta é uma das razões pelas quais os compostos seletivos não se traduziram automaticamente em perfis clínicos limpos na dor, na doença psiquiátrica ou nas perturbações inflamatórias, apesar da elevada necessidade não satisfeita. A dor crónica, só por si, afeta quase 1 em cada 5 adultos nos Estados Unidos, segundo a National Academies, mas a farmacologia canabinoide não produziu uma solução universalmente fiável baseada em recetores.
Visto desta forma, CB1 e CB2 são melhor entendidos como nós de controlo inseridos em circuitos lipídicos, imunitários e sinápticos. A sua capacidade de serem alvos terapêuticos é real. A sua capacidade de surpreender também.
CB1 como alvo terapêutico: promessa, limitações e o problema dos efeitos secundários centrais
CB1 continua a ser um dos alvos mais tentadores na neurofarmacologia, porque ocupa um ponto estratégico de controlo na sinalização sináptica. O trabalho de farmacologia dos recetores de Allyn Howlett estabeleceu que os efeitos do cannabis não eram um fenómeno inespecífico da membrana, mas sim biologia mediada por recetores, e a posterior descoberta da anandamida por Raphael Mechoulam e Lumír Hanuš deu a esse recetor um sistema de sinalização endógeno. Essa história é importante. Um recetor que ajuda a modular a libertação de neurotransmissores em circuitos da dor, circuitos da alimentação, vias do stress e redes de recompensa parece uma ferramenta plausível para várias perturbações difíceis em simultâneo. Mas também cria uma armadilha: quando um recetor está integrado em tantas funções cerebrais essenciais, a manipulação ampla raramente permanece confinada ao efeito desejado.
Esse problema não é académico. A Organização Mundial de Saúde estima que 200 milhões de pessoas usaram cannabis em 2019, cerca de 4% da população global com idades entre os 15 e os 64 anos, pelo que a farmacologia do CB1 já está a operar à escala populacional através dos cannabinoid de origem vegetal, especialmente delta-9-THC. Ainda assim, os medicamentos cannabinoid aprovados continuam a ser poucos: a FDA dos EUA afirma que, em 2025, aprovou um produto medicamento derivado de cannabis e três produtos medicamentosos relacionados com cannabis. A diferença entre a exposição generalizada e as terapêuticas aprovadas limitadas diz algo importante sobre o CB1. A biologia é convincente. A concretização farmacológica é difícil.
Fundamentação terapêutica na dor, apetite e neuropsiquiatria
O interesse começa pelo local de ação do CB1 e pelo que ele faz. O CB1 é altamente expresso em muitos terminais pré-sinápticos no sistema nervoso central, onde os endocannabinoid são frequentemente produzidos “on demand” nas células pós-sinápticas e se deslocam no sentido retrógrado através da sinapse para suprimir a libertação de neurotransmissores. Este sistema retrógrado pode atenuar glutamato, GABA e outros fluxos de sinalização, dependendo do tipo celular e do circuito. Para a dor, essa é uma fundamentação óbvia: se quase 1 em cada 5 adultos nos Estados Unidos vive com dor crónica, como referiu a National Academies em 2017, um recetor capaz de reduzir a transmissão nociceptiva e de alterar o componente afetivo da dor vai inevitavelmente atrair atenção.
Mas o CB1 não é um simples interruptor analgésico. Em alguns circuitos, a supressão da libertação inibitória de GABA pode desinibir neurónios a jusante, em vez de os acalmar. Noutros, predomina a supressão glutamatérgica. O efeito depende da sinapse, do estado da rede e do ligando. Os endocannabinoid, como a anandamida e o 2-AG, são sinais breves, gerados localmente. Os phytocannabinoid e os agonistas sintéticos chegam com cinéticas diferentes, eficácia diferente e, muitas vezes, com um envolvimento recetorial muito mais amplo ao longo do tempo. Essa distinção ajuda a explicar por que motivo “a ativação do CB1 reduz a dor” é uma formulação demasiado grosseira para orientar um desenho clínico seguro.
O apetite é uma segunda grande fundamentação. A sinalização do CB1 promove a ingestão alimentar e interage com circuitos hipotalâmicos e mesolímbicos que ligam a homeostase energética à recompensa. Isto tornou o antagonismo do CB1 uma estratégia anti-obesidade apelativa e a agonismo do CB1, ou o seu reforço indireto, uma via potencial para caquexia ou perda de apetite. A lógica não era irracional. Estava enraizada na fisiologia básica. Contudo, o apetite é inseparável do humor, da saliência e do processamento do stress, todos eles também moldados pelo CB1.
É na neuropsiquiatria que o recetor se torna mais interessante e mais perigoso. O CB1 modula circuitos relevantes para a dopamina, a inibição cortical, a plasticidade hipocampal, a aprendizagem do medo e a responsividade ao stress. Isso confere-lhe relevância teórica para perturbações de ansiedade, condições relacionadas com trauma, depressão, perturbações por uso de substâncias e doença psicótica. O American Journal of Psychiatry argumentou, em 2025, que a sinalização enviesada do CB1 é uma estratégia plausível para a esquizofrenia, uma perturbação que afeta cerca de 24 milhões de pessoas em todo o mundo, segundo a WHO. A importância desse artigo não é a de demonstrar que um medicamento dirigido ao CB1 para a esquizofrenia está pronto. É a de mostrar que a área evoluiu para lá do agonismo e antagonismo grosseiros, em direção a uma farmacologia seletiva por via, inspirada no quadro mais amplo dos GPCR.
Porque é que o direcionamento do CB1 é cientificamente apelativo, mas clinicamente difícil
O CB1 é cientificamente apelativo exatamente pela mesma razão pela qual é clinicamente difícil: é um núcleo de sinalização denso. No nível canónico, o CB1 acopla-se sobretudo a proteínas Gi/o, reduzindo a atividade da adenilato ciclase, modulando canais iónicos e suprimindo a libertação de neurotransmissores. Só isso já bastaria para o tornar importante. Mas também recruta beta-arrestinas, sofre dessensibilização e internalização, e pode ser moldado por moduladores alostéricos e por estados conformacionais específicos de ligando. A revisão estrutural da Frontiers in Chemical Biology de 2026 torna este ponto claro para o CB1 e o CB2: a seletividade e a eficácia dos ligandos decorrem de diferenças estruturais ao nível do recetor que afetam a ligação, a saída de sinalização e a regulação do recetor. Cannabinoid com aspeto semelhante podem, portanto, produzir farmacologia significativamente diferente.
Isto não é apenas um pormenor de química medicinal. Vai diretamente aos efeitos secundários. Um ligando que impulsione fortemente uma conformação ativa pode provocar maior internalização do recetor, mais tolerância ou um perfil comportamental diferente de um ligando que estabilize outro estado. Um estudo indexado no PubMed de 2025/2026 sobre seletividade de subtipos argumentou ainda que a seletividade dos endocannabinoid é dinâmica, e não um evento fixo de “chave-fechadura”, reforçando a ideia de que o comportamento do recetor muda consoante o contexto conformacional. Os desenvolvedores de medicamentos não enfrentam um único alvo CB1. Enfrentam um conjunto em movimento.
É por isso que agonistas e antagonistas diretos do CB1 têm desapontado tantas vezes. Um agonismo central forte pode prejudicar a memória, a atenção, o desempenho psicomotor e a regulação da ansiedade, ao mesmo tempo que ativa vias de recompensa de forma a complicar o risco de dependência. Um antagonismo central forte pode reduzir o apetite, sim, mas também pode interferir com os mesmos circuitos de amortecimento do stress e afetivos que os endocannabinoid endógenos normalmente estabilizam. O CB1 está profundamente envolvido na cognição, recompensa e regulação do stress. Qualquer medicamento que force demasiado esse sistema numa direção deve ser considerado de risco até prova em contrário.
A biologia de sistemas sustenta esta cautela. Uma análise integrativa de redes de 2025/2026 identificou o CB1 e o CB2 como nós altamente influentes no sistema endocannabinoid e ligou a sinalização do recetor a vias metabólicas mais vastas. Esse achado argumenta contra a simplificação centrada apenas no recetor. Alterar o CB1 não desloca apenas uma sinapse. Pode propagar-se por redes endócrinas, imunitárias, metabólicas e comportamentais.
Lições das estratégias com ação central versus restrição periférica
A lição mais clara da primeira era do desenvolvimento de medicamentos dirigidos ao CB1 é que a exposição central é, muitas vezes, o ponto em que eficácia e toxicidade se tornam inseparáveis. Agonistas do CB1 com ação central podem envolver circuitos da dor e do apetite, mas a mesma penetração cerebral aumenta a probabilidade de efeitos semelhantes à intoxicação, sedação, perturbação cognitiva e eventos adversos psiquiátricos. Os antagonistas do CB1 com ação central ilustram o modo de falha oposto: podem atingir objetivos metabólicos, mas produzir efeitos inaceitáveis no humor e na ansiedade, porque também bloqueiam o tónus cannabinoid endógeno em circuitos límbicos.
Essa história é a razão pela qual os ligandos do CB1 com restrição periférica se tornaram apelativos. A ideia é simples: se alguns benefícios terapêuticos dependem do CB1 em tecidos periféricos, como aferentes primários, intestino, fígado, tecido adiposo ou outros locais fora do SNC, então manter o medicamento fora do cérebro poderá preservar efeitos úteis enquanto reduz os efeitos adversos centrais. Para a dor, isto pode significar atuar sobre mecanismos periféricos nociceptivos. Para a doença metabólica, pode significar reduzir a lipogénese ou alterar vias relacionadas com a glicose sem perturbar o humor ou a cognição.
A lógica é sólida, mas não é uma solução mágica. Primeiro, a biologia do CB1 periférico e central não é nitidamente separável em estados de doença. Dor crónica, alimentação e stress envolvem circuitos de retroalimentação corpo-cérebro. Segundo, a exclusão pela barreira hematoencefálica é um problema quantitativo, não binário. Pequenas quantidades de penetração no SNC podem continuar a ser relevantes, sobretudo com administração crónica. Terceiro, mesmo um ligando do CB1 verdadeiramente periférico pode falhar clinicamente se a doença-alvo depender fortemente de circuitos centrais ou se vias compensatórias reduzirem a eficácia.
A estratégia mais recente não é apenas “manter-se fora do cérebro”. É “moldar o sinal”. Isso inclui agonistas parciais, antagonistas neutros em vez de agonistas inversos, moduladores alostéricos e ligandos enviesados que favorecem vias downstream selecionadas em detrimento de outras. A discussão do American Journal of Psychiatry de 2025 sobre a sinalização enviesada do CB1 na esquizofrenia enquadra-se aqui. Se um ligando puder preservar sinalização terapêutica selecionada enquanto reduz o recrutamento de beta-arrestina, a dessensibilização do recetor ou os efeitos indesejados do circuito, então o CB1 poderá tornar-se mais manejável. Poderá. Ainda não há garantia de que o enviesamento de via observado em sistemas celulares se traduza de forma limpa nas janelas terapêuticas humanas.
Assim, a visão atual do CB1 como alvo terapêutico não é nem desvalorizadora nem romântica. O CB1 é apelativo porque regula o ganho sináptico precisamente nos tipos de circuitos que os clínicos desejam influenciar. É difícil porque esses circuitos também regulam a memória, o afeto, a motivação e a resiliência ao stress. Ligandos com restrição periférica, estratégias de sinalização seletiva e uma melhor compreensão estrutural melhoraram a lógica do desenho. Não eliminaram o problema central. Um recetor tão central para a função cerebral raramente tolera farmacologia grosseira.
CB2 como alvo terapêutico: neuroinflamação, neurodegeneração e modulação imunitária
O CB2 tornou-se um dos alvos mais atractivos na farmacologia dos cannabinoid por uma razão simples: parece oferecer acesso à biologia imunitária e glial com menor risco de intoxicação, défice de memória e potencial de abuso classicamente associados a uma forte activação do CB1. Esse atractivo é importante muito para além do mapeamento académico de recetores. A World Health Organization estimou que 200 milhões de pessoas usaram cannabis em 2019, ou 4% da população mundial com idades entre os 15 e os 64 anos, e a U.S. FDA ainda lista, em 2025, apenas um medicamento derivado de cannabis e três medicamentos relacionados com cannabis aprovados. O interesse é elevado. A tradução é difícil.
A antiga formulação resumida dizia que o CB1 é o recetor cerebral e o CB2 é o recetor imunitário. Isso é agora demasiado simplista para ser útil. A revisão de 2026 na Frontiers in Behavioral Neuroscience, explicitamente enquadrada como “an update over the last 3 years”, trata o CB2 como relevante para perturbações do sistema nervoso central precisamente porque participa em mecanismos neuroinflamatórios e neurodegenerativos, incluindo activação microglial, sinalização de citocinas e comunicação neurónio-glia. A alteração fundamental não é o CB2 ter subitamente passado a ser abundante em todo o cérebro. Não passou. A alteração é que a baixa expressão basal em muitas regiões do SNC não significa irrelevância funcional, sobretudo quando estados de doença podem alterar a expressão, o tráfego e a sinalização dos recetores.
Porque é que o CB2 é frequentemente visto como um alvo mais seguro
O argumento de segurança para o CB2 começa pela distribuição, mas não termina aí. O CB2 é enriquecido em células imunitárias e é frequentemente sobrerregulado em microglia activada, macrófagos infiltrados e outras populações celulares inflamatórias. Em contraste, o CB1 é densamente expresso em terminais pré-sinápticos no córtex, hipocampo, gânglios da base e cerebelo, onde molda fortemente a libertação de neurotransmissores. Essa diferença anatómica é importante porque um ligando que interaja sobretudo com o CB2 tem menor probabilidade de reproduzir o perfil psicoactivo clássico associado ao CB1.
Menor probabilidade não é o mesmo que impossibilidade. A selectividade é um alvo em movimento. A revisão de 2026 na Frontiers in Chemical Biology sublinha que a selectividade entre “CB1 and CB2” depende de diferenças estruturais que alteram a ligação, a eficácia e a regulação dos recetores, e um estudo recente indexado na PubMed sobre selectividade de subtipos argumenta que a discriminação endocannabinoid entre subtipos de recetores reflecte dinâmica conformacional e não um modelo fixo de chave-fechadura. Na prática, isto significa que um composto pode parecer selectivo para CB2 num ensaio e comportar-se de forma menos limpa noutro, sobretudo quando a densidade de recetores, o ambiente de membrana e os transdutores a jusante diferem. O mesmo ligando pode enviesar a sinalização por G-proteína num tipo celular, recrutar beta-arrestinas noutro e apresentar diferentes cinéticas de dessensibilização ou internalização em condições inflamatórias.
Essa complexidade mecanística, na verdade, reforça o interesse em prosseguir o CB2 com cuidado. Se o objectivo terapêutico é reduzir a produção de citocinas pela microglia, limitar o recrutamento de células imunitárias periféricas ou alterar o stress metabólico glial sem perturbar fortemente a transmissão sináptica cortical, então o CB2 continua a ser o recetor mais plausível para começar. A sinalização enviesada do CB1, como defendido no artigo de 2025 do American Journal of Psychiatry sobre esquizofrenia, pode eventualmente alargar a janela terapêutica para fármacos dirigidos ao CB1. Mas esse é um argumento a favor de uma farmacologia do CB1 mais inteligente, não uma razão para ignorar a menor carga de psicoactividade das abordagens centradas no CB2.
Mais seguro, porém, não deve ser confundido com eficaz. Um fármaco pode evitar intoxicação óbvia e ainda assim falhar porque o recetor é expresso de forma demasiado escassa, demasiado variável ou demasiado transitória no processo de doença para gerar um efeito clínico relevante. Isso aconteceu repetidamente no desenvolvimento de fármacos para neuroinflamação, e não apenas na ciência do cannabis.
Evidência e limitações em modelos de doença do SNC
A revisão da Frontiers in Behavioral Neuroscience explica por que motivo o CB2 continua a surgir em modelos de doença de Alzheimer, doença de Parkinson, esclerose múltipla, traumatismo cranioencefálico, acidente vascular cerebral e dor neuropática: todas estas condições incluem sinalização inflamatória, reactividade glial, stress oxidativo e lesão tecidular progressiva. Em muitos estudos em roedores, agonistas do CB2 reduzem mediadores pró-inflamatórios como TNF-α, IL-1β e IL-6, deslocam a microglia para fenótipos menos lesivos ou atenuam a infiltração de leucócitos. Por vezes, isso vem acompanhado de melhoria do comportamento motor, redução do tamanho da lesão ou preservação de marcadores sinápticos. Esses são sinais reais, não artefactos a descartar.
Mas o conjunto de dados é irregular. Os efeitos dependem frequentemente da janela temporal. Um agonista do CB2 administrado durante a fase inflamatória inicial após uma lesão pode fazer algo muito diferente do mesmo agonista administrado durante uma degeneração crónica. A dose também importa, tal como a classe do ligando. Os endocannabinoid como 2-AG e anandamida não são intercambiáveis com phytocannabinoid nem com agonistas sintéticos com preferência para CB2. As suas concentrações locais, degradação metabólica e acções fora do alvo diferem. O mesmo acontece com as preferências de via.
Os próprios modelos de doença são outra limitação. Um rato transgénico de Alzheimer com patologia amiloide e microglia reactiva não é a doença de Alzheimer num humano de 74 anos com falência vascular, inflamatória e proteostática combinada. A encefalomielite autoimune experimental é útil para a biologia da esclerose múltipla, mas captura apenas parte da condição humana. Assim, os resultados pré-clínicos positivos estabelecem plausibilidade, não prova.
Há também um problema persistente de medição na biologia do CB2. A especificidade de anticorpos tem sido, há muito, um ponto fraco, tornando as alegações de localização do recetor mais difíceis de confiar do que muitos artigos sugerem. Os baixos níveis de expressão no cérebro saudável, a expressão induzível em doença e as diferenças entre espécies complicam a interpretação. Alguns resultados relatados de “CB2 neuronal” podem ser reais em contextos específicos, mas as alegações amplas de expressão neuronal constitutiva e disseminada do CB2 continuam menos sólidas do que o campo sugeria anteriormente.
É aqui que a sinalização importa tanto quanto a anatomia. O CB2 é um GPCR acoplado a Gi/o, mas essa designação esconde variações importantes: inibição da adenylyl cyclase, modulação de vias MAPK, efeitos em canais iónicos, recrutamento de beta-arrestina, dessensibilização do recetor e internalização podem alterar o resultado biológico líquido. Um ligando que reduz a libertação de citocinas em microglia em cultura pode falhar in vivo se dessensibilizar rapidamente os recetores ou se desencadear um programa dominado por arrestina pouco útil. Menor risco de intoxicação não resolve estes problemas farmacológicos.
O que a literatura recente sugere para terapias futuras
A literatura recente afasta-se da antiga procura de um único “agonista do CB2 para a inflamação cerebral” e avança para uma estratégia mais selectiva e dependente do contexto. A revisão estrutural de 2026 e o artigo de 2025/2026 sobre selectividade de subtipo apoiam ambos a mesma lição: o direccionamento do recetor dependerá do estado conformacional, do perfil de eficácia e do contexto tecidular, e não apenas da afinidade nominal por um subtipo. O desenho de fármacos está a evoluir para ligandos com selectividade funcional, e não apenas de ligação.
Isto provavelmente significa três coisas. Primeiro, os futuros fármacos do CB2 terão de apresentar perfis de sinalização melhores, incluindo enviesamento entre G-proteína e beta-arrestina, tempo de residência no recetor e efeitos específicos por tipo celular em sistemas humanos, e não apenas em linhas celulares imortalizadas de ensaio. Segundo, a estratificação da doença será importante. Uma terapêutica dirigida ao CB2 pode funcionar melhor quando a amplificação inflamatória é um motor principal da patologia, e não apenas um subproduto. Terceiro, a ideia de combinação é inevitável. Uma análise de rede integrativa publicada num estudo recente indexado na PubMed identificou o CB1 e o CB2 como nós influentes em vias mais amplas de endocannabinoid e metabólicas, o que está em conformidade com a realidade clínica da doença do SNC: inflamação, stress mitocondrial, sinalização lipídica e disfunção sináptica estão interligados.
Em condições inflamatórias específicas, a modulação do CB2 pode reduzir a sinalização imunitária danosa, evitando em grande parte a carga psicoativa ligada ao CB1.Preliminary evidence
Isso não faz do CB2 um beco sem saída. Faz dele um alvo que provavelmente pertence à farmacologia de precisão, e não à mitologia dos recetores. O argumento actual mais forte não é “a activação do CB2 trata a neurodegeneração”. É mais restrito: em condições inflamatórias específicas, em populações celulares específicas, com as propriedades corretas do ligando, a modulação do CB2 pode reduzir a sinalização imunitária lesiva enquanto evita grande parte do fardo psicoactivo associado ao CB1. Essa é uma hipótese terapêutica credível. Ainda não é uma regra clinicamente validada.
Compostos de cannabis em CB1 e CB2: THC, CBD, canabinoides minoritários e ligandos sintéticos
A cannabis não actua através de uma única substância química nem de um único mecanismo receptor. Esse ponto parece óbvio, mas o debate público ainda reduz a farmacologia a uma associação falsa: THC equivale a CB1, CBD equivale a CB2. A literatura não apoia esse atalho. A cannabis contém muitos fitocanabinoides, além de terpenos, flavonoides e outros constituintes, enquanto o organismo produz os seus próprios endocanabinoides, como a anandamida e o 2-arachidonoilglicerol, identificados através de trabalhos associados a Raphael Mechoulam, Lumír Hanuš e outros. Para além disso, a maior parte do que os cientistas sabem sobre activação de receptores, eficácia, selectividade, dessensibilização e enviesamento de sinalização provém de ligandos sintéticos concebidos como sondas ou como compostos líderes de desenvolvimento farmacológico, e não apenas de compostos da planta.
Essa distinção é importante porque CB1 e CB2 não são fechaduras passivas à espera de uma chave canabinoide. São receptores acoplados à proteína G da classe A, com múltiplos estados conformacionais, preferências de acoplamento e destinos regulatórios. Uma revisão estrutural de 2026 em Frontiers in Chemical Biology deixa o ponto claro: a selectividade dos ligandos em CB1 e CB2 depende de diferenças estruturais ao nível do receptor que alteram a ligação, a eficácia e a regulação do receptor, e não apenas do facto de uma molécula “atingir” um receptor ou outro. Um estudo recente indexado no PubMed sobre selectividade entre subtipos argumenta igualmente que a preferência dos endocanabinoides por CB1 versus CB2 resulta de dinâmicas conformacionais, e não de um simples encaixe chave-fechadura. Por isso, quando dois canabinoides parecem quimicamente semelhantes, mas produzem efeitos diferentes no cérebro, no tecido imunitário ou nos circuitos da dor, isso não é um detalhe secundário. É a farmacologia central.
Tendo em conta que a Organização Mundial da Saúde estimou que 200 milhões de pessoas consumiram cannabis em 2019, cerca de 4% da população mundial entre os 15 e os 64 anos, estas distinções não são trivia académica. Moldam a intoxicação, as alegações terapêuticas, a segurança e a razão pela qual alguns fármacos direccionados a receptores têm sucesso enquanto outros falham.
Como o THC interage com CB1 e contribui para os efeitos psicoactivos
Delta-9-tetra-hidrocanabinol, ou THC, continua a ser o composto central para compreender a intoxicação por cannabis porque interage directamente com CB1 no sistema nervoso central. Essa é a explicação mais clara ao nível do receptor para os efeitos psicoactivos característicos da cannabis, ainda que “psicoactivo” abranja vários desfechos separáveis: alteração da percepção temporal, recompensa, perturbação da memória, ansiedade em alguns utilizadores e prejuízo da coordenação motora. O trabalho de farmacologia de receptores de Allyn Howlett ajudou a estabelecer este quadro centrado em CB1, e ele continua a ser fundamental.
Mas “THC activa CB1” é apenas o início. O THC não é um interruptor perfeito. Em geral, é descrito como um agonista parcial em CB1, o que significa que a sua eficácia depende da densidade de receptores, do contexto tecidular e do ensaio de sinalização utilizado. Em regiões ricas em CB1, como o córtex, o hipocampo, os gânglios da base e o cerebelo, isso pode ser suficiente para produzir efeitos centrais intensos. Ao nível sináptico, CB1 encontra-se frequentemente na pré-sinapse, onde a activação suprime a libertação de neurotransmissores. Isto significa que o THC pode reduzir a libertação de glutamato num circuito, a libertação de GABA noutro, e produzir efeitos opostos ao nível da rede, dependendo de quais as células que expressam o receptor e em que momento.
É por isso que a localização do receptor não pode ser reduzida a “CB1 no cérebro”. CB1 é abundante no cérebro, sim, mas não de forma uniforme, não de forma estática e não de forma funcionalmente idêntica entre tipos neuronais. O resultado é que os efeitos do THC são dependentes do circuito. A euforia e a reforçação fazem parte do quadro. Tal como a perturbação da memória de curto prazo e a sinalização relevante para psicose em pessoas susceptíveis.
Esse último ponto tornou-se mais importante, e não menos. Um artigo de 2025 no American Journal of Psychiatry defendeu que a sinalização enviesada de CB1 é uma estratégia terapêutica plausível na esquizofrenia, precisamente porque a biologia de CB1 não pode ser entendida como uma activação receptorial unidimensional. A esquizofrenia afecta cerca de 24 milhões de pessoas em todo o mundo, segundo a WHO, e qualquer discussão séria sobre THC em CB1 tem de considerar o facto de a mesma família de receptores implicada na intoxicação estar também a ser estudada através de agonismo enviesado, recrutamento de beta-arrestina e desenho farmacológico selectivo por via. O THC pode interagir com a sinalização acoplada a Gi/o, mas CB1 também sofre fosforilação, dessensibilização e internalização, com consequências para a tolerância e a resposta a jusante. A dose e o timing importam. A exposição repetida altera o sistema.
Os ligandos sintéticos de CB1 deixaram isto claro há muito tempo. Compostos como CP55,940 e WIN55,212-2 são agonistas potentes de investigação que frequentemente produzem respostas receptorais mais intensas ou mais nítidas do que o THC em ensaios laboratoriais. O Rimonabant, um inverso agonista de CB1, mostrou a direcção oposta: bloquear ou empurrar CB1 para abaixo da sua actividade basal podia reduzir o apetite e o peso corporal, mas os efeitos adversos psiquiátricos deitaram o fármaco por terra. Essa história clínica é um aviso. A focalização selectiva em CB1 pode ser farmacologicamente elegante e ainda assim medicamente decepcionante.
Porque é que o CBD não se enquadra numa história simples de agonista CB1/CB2
O CBD é o canabinoide que mais frequentemente é reduzido a mito. É rotineiramente apresentado como “o não intoxicante que actua através de CB2” ou como o oposto directo do THC ao nível do receptor. Nenhuma destas afirmações é exacta.
| Classe de composto ou exemplo | Como o artigo descreve o comportamento do recetor | Conclusão principal |
|---|---|---|
| THC | Agonista parcial de CB1 e CB2; central para a intoxicação através de CB1 | O envolvimento de CB1 ajuda a explicar os efeitos psicoativos, mas o resultado continua dependente do contexto |
| CBD | Não é um agonista simples de CB1 ou CB2; pode atuar de forma alostérica ou indireta | O CBD não é uma imagem especular simples do THC em CB2 |
| Canabinoides menores | Os efeitos relatados variam consoante o ensaio e a concentração | Não generalizar o comportamento do recetor de um canabinoide para todos |
| Ligandos sintéticos | Frequentemente usados para definir eficácia, seletividade e enviesamento | Os compostos sonda podem comportar-se de forma muito diferente dos canabinoides da planta |
O CBD não se comporta como um agonista simples de CB1 ou CB2, como o THC faz em CB1. A sua afinidade pelos sítios ortostéricos de CB1 e CB2 é relativamente baixa, e grande parte da sua farmacologia parece envolver acções indirectas, alostéricas ou em receptores não canabinoides. Dependendo do sistema estudado, foi descrito como modulador alostérico negativo em CB1, o que poderia alterar a forma como o THC ou os endocanabinoides sinalizam sem simplesmente ligar ou desligar o receptor. Essa distinção é crucial. Modificar a forma do receptor e a eficiência da sinalização não é o mesmo que agonismo clássico.
O CBD também interage com alvos para além de CB1 e CB2, incluindo TRPV1, 5-HT1A, PPAR-gamma, vias relacionadas com adenosina e enzimas envolvidas no tónus endocanabinoide. Alguns efeitos podem reflectir alteração da sinalização da anandamida em vez de estimulação directa do receptor. Isso ajuda a explicar porque é que o perfil clínico do CBD não se alinha de forma limpa com a antiga abreviatura receptorial. A FDA dos EUA refere que aprovou um produto medicamentoso derivado de cannabis e três produtos medicinais relacionados com cannabis até 2025; o uso médico mais bem estabelecido do CBD situa-se na epilepsia, uma condição que afecta cerca de 50 milhões de pessoas em todo o mundo, e essa via terapêutica não pode ser explicada por um modelo caricatural de “o CBD liga-se a CB2 e reduz a inflamação”.
Mesmo em relação a CB2, o quadro é mais amplo do que a narrativa periférica antiga. Uma revisão de 2026 em Frontiers in Behavioral Neuroscience descreveu “uma actualização ao longo dos últimos 3 anos” em que a sinalização de CB2 ganhou atenção em perturbações do sistema nervoso central através de ligações a mecanismos neuroinflamatórios e neurodegenerativos. Isso não significa que o CBD seja simplesmente um fármaco de CB2. Significa que o próprio receptor tem uma pegada funcional maior do que a história pública sugere.
A importância de separar os compostos da planta dos mitos sobre receptores
Os fitocanabinoides não são intercambiáveis, e também não são intercambiáveis com ligandos sintéticos. Canabinoides minoritários como canabigerol (CBG), canabinol (CBN), tetrahidrocanabivarina (THCV) e canabicromeno (CBC) foram descritos como interagindo com CB1 e CB2 de formas que variam consoante o ensaio e a concentração. Alguns mostram agonismo fraco, outros agonismo parcial, outros antagonismo em determinadas condições, e outros actividade relevante em alvos não canabinoides. THCV é um bom exemplo: foi descrito como antagonista de CB1 ou antagonista neutro em doses baixas em alguns sistemas e como agonista parcial em doses mais elevadas noutros. Isso não é inconsistencia por si só. Reflecte farmacologia receptorial dependente do contexto.
As ferramentas de investigação tornam este ponto mais nítido. Endocanabinoides, fitocanabinoides e ligandos sintéticos podem todos activar “CB1 e CB2”, mas não estabilizam as mesmas conformações receptorais nem produzem o mesmo grau de enviesamento entre proteínas G e beta-arrestinas. A revisão estrutural de 2026 e as análises de rede recentes indexadas no PubMed afastam o campo da mitologia dos receptores e aproximam-no da farmacologia de sistemas. Os receptores existem dentro de redes de sinalização específicas de cada célula, vias metabólicas e maquinaria de tráfego intracelular. O perfil clínico de um ligando emerge desse contexto completo, e não de uma etiqueta como “CB2 anti-inflamatório” ou “CBD equilibra o THC”.
Para uma wiki sobre cannabis, essa é a linha mais importante: o THC tem relevância primária para a intoxicação mediada por CB1, mas a farmacologia da cannabis não termina aí, e o CBD não é uma simples imagem espelhada do THC em CB2. Os compostos da planta iniciam a história. O estado do receptor, a distribuição tecidular, a eficácia, o enviesamento e a regulação decidem como ela termina.
Esquizofrenia, risco de psicose e o que a biologia do CB1 implica e não implica
Porque é que a esquizofrenia entra na conversa sobre o receptor
A esquizofrenia é relevante aqui porque o CB1 não é apenas mais um receptor numa longa lista de alvos. É um dos receptores acoplados à proteína G mais abundantes no cérebro, localizado em terminações pré-sinápticas onde pode atenuar a libertação de transmissores e remodelar a atividade de rede em circuitos já implicados na psicose: córtex, hipocampo, estriado, amígdala. Quando um receptor está tão próximo do controlo de circuitos relacionados com glutamato, GABA, dopamina e responsividade ao stresse, os investigadores em psiquiatria prestam atenção.
A escala de saúde pública torna esta questão mais do que académica. A Organização Mundial da Saúde estima que o cannabis foi utilizado por 200 milhões de pessoas em 2019, ou 4% da população mundial com idades entre os 15 e os 64 anos. A WHO estima também que a esquizofrenia afeta cerca de 24 milhões de pessoas em todo o mundo, aproximadamente 1 em cada 300 pessoas. Não se trata de populações pequenas e isoladas. Sobrepõem-se em clínicas reais, serviços de urgência e estudos de coorte longitudinais. Qualquer biologia de recetor que possa esclarecer o risco de psicose, o desenvolvimento de fármacos antipsicóticos, ou ambos, merece um escrutínio sério.
Esse é o contexto translacional do artigo de 2025 no American Journal of Psychiatry que argumenta que a sinalização enviesada do CB1 é uma estratégia terapêutica plausível para a esquizofrenia. A ideia central é fácil de enunciar e fácil de deturpar. Fácil de enunciar: o CB1 não se comporta como um simples interruptor de ligar/desligar, e diferentes ligandos podem estabilizar diferentes conformações do recetor, desviando a sinalização para, ou afastando-a de, vias intracelulares específicas. Fácil de deturpar: se o CB1 é relevante para a investigação do tratamento da esquizofrenia, alguns leitores saltam para a afirmação falsa de que “a ativação dos cannabinoid ajuda a esquizofrenia” ou de que o consumo de cannabis é, por isso, medicinal por defeito. O artigo não apoia esse salto.
Historicamente, a ciência dos cannabinoid ficou presa a abreviações simplistas. O CB1 foi apresentado como o “receptor do cérebro”, o CB2 como o “receptor imunitário”, e os efeitos psicoativos foram atribuídos quase exclusivamente à ocupação do CB1. Essa formulação sempre foi incompleta e, em 2025, é claramente demasiado grosseira. O trabalho fundamental de Allyn Howlett estabeleceu a farmacologia dos receptores cannabinoid, enquanto Mechoulam e Hanus ajudaram a definir a biologia do endocannabinoid através da anandamida. Desde então, a farmacologia dos recetores passou da presença do recetor para o estado do recetor. Essa mudança é importante para a esquizofrenia porque a fisiopatologia não é causada apenas pela expressão do recetor. Surge da temporalidade, do tipo celular, do contexto do circuito e da resposta intracelular.
A literatura mais recente reforça esse ponto. Uma revisão de 2026 na Frontiers in Chemical Biology explica que as diferenças estruturais entre CB1 e CB2 moldam a ligação do ligando, a eficácia, a sinalização e a regulação do recetor. Um estudo indexado no PubMed de 2025/2026 sobre seletividade de subtipo argumenta que a seletividade dos endocannabinoid pode resultar de dinâmicas conformacionais e não de um modelo estático de fechadura e chave. Outro artigo de análise em rede de 2025/2026 identifica CB1 e CB2 como nós altamente influentes em todo o sistema endocannabinoid, ligados a vias metabólicas e de sinalização, e não a caixas de recetores isoladas. Para a investigação da esquizofrenia, isto significa que se deve deixar de perguntar apenas “esta substância atua no CB1?” e começar a perguntar “que estados do CB1, em que células, sob que condições de exposição?”
É daí que vem a seriedade clínica. Não de conversa sobre recetores ao nível de slogans. Mas de precisão mecanística.
Sinalização enviesada versus ativação bruta do recetor
A ativação bruta do CB1 é um modelo pobre para o raciocínio terapêutico. Reduz demasiadas variáveis a uma só palavra: ativação. Ligandos endógenos como a anandamida e o 2-AG, fitocannabinoid como o delta-9-THC e agonistas sintéticos não se comportam de forma idêntica no CB1, mesmo quando todos contam como ligandos. Podem diferir na afinidade, na eficácia, no tempo de residência, na exposição regional e no enviesamento de via. Também podem diferir na intensidade com que recrutam proteínas Gi/o em comparação com beta-arrestinas, na rapidez com que desencadeiam dessensibilização e na extensão da internalização do recetor após doses repetidas.
Essa distinção é central para o argumento sobre a esquizofrenia no AJP. Um ligando que inclina o CB1 para um programa intracelular pode não se parecer com um ligando que impulsiona uma ativação ampla do recetor em muitas regiões do cérebro. Na farmacologia dos GPCR, essa diferença pode separar um fármaco útil de um intolerável. O CB1 é especialmente sensível a este problema porque o mesmo recetor pode reduzir a transmissão excitatória numa sinapse, suprimir a transmissão inibitória noutra e alterar a plasticidade a jusante de formas que não são funcionalmente equivalentes. Pequenas diferenças moleculares podem produzir grandes consequências ao nível dos sistemas.
THC é o exemplo óbvio de porque “alvo CB1” não é igual a “terapia CB1”. O THC é um fitocannabinoid com propriedades de agonista parcial no CB1, mas os efeitos vividos e clínicos do THC não são uma leitura pura de um único ramo de sinalização. A dose importa. A idade na exposição importa. A exposição repetida importa. O mesmo acontece com a proporção de THC face a outros constituintes e com a vulnerabilidade psiquiátrica da pessoa. Efeitos agudos de intoxicação, ansiedade, alteração percetiva e sintomas psicóticos transitórios em alguns utilizadores não nos dizem que todas as terapêuticas dirigidas ao CB1 estão condenadas. Dizem-nos, sim, que a estimulação indiscriminada do CB1 não é um atalho para o tratamento antipsicótico.
A agonismo enviesado é atrativo precisamente porque tenta evitar essa falta de precisão. A esperança terapêutica não é “mais CB1”. É “o perfil de sinalização correto do CB1”. Isso pode significar favorecer vias de sinalização associadas à estabilização de circuitos, limitando ao mesmo tempo vias ligadas a efeitos cognitivos ou psicotomiméticos adversos, tolerância ou downregulation do recetor. Se isto pode ser alcançado em doentes continua a ser uma questão em aberto. O conceito é plausível. Não é um sucesso clínico comprovado.
O CB2 também pertence a este quadro, embora não como uma solução simplista. A revisão de 2026 na Frontiers in Behavioral Neuroscience descreve “uma atualização dos últimos 3 anos” em que a sinalização do CB2 ganhou atenção nos distúrbios do sistema nervoso central devido a ligações neuroinflamatórias e neurodegenerativas. Isso enfraquece a velha afirmação de que o CB2 é irrelevante para a doença cerebral. Mas não resgata a antiga dicotomia ao invertê-la. A esquizofrenia não é apenas inflamação, e o CB2 não é apenas um botão anti-inflamatório. Qualquer modelo translacional sério tem de considerar em conjunto as interações neurónio-glia, a sinalização imunitária e a disfunção dos circuitos.
O que não inferir
- Não é prova de benefício O facto de o CB1 ser um alvo farmacológico não significa que o consumo de cannabis proteja contra a esquizofrenia ou a psicose.
- Não refuta o risco Estudos mecanísticos sobre recetores não anulam a preocupação epidemiológica relativa à exposição a THC elevado em pessoas vulneráveis.
- Não são intercambiáveis Endocanabinoides, fitocanabinoides e ligandos sintéticos não devem ser tratados como o mesmo tipo de exposição.
- Não é aprovação indiscriminada As aprovações existentes da FDA não resolvem questões de segurança psiquiátrica ou de eficácia para o uso amplo de canabinoides.
O que os leitores não devem inferir a partir dos estudos mecanísticos
Em primeiro lugar, os leitores não devem inferir que, porque o CB1 é um alvo terapêutico, o consumo de cannabis é, por isso, protetor contra a esquizofrenia ou a psicose. A lógica alvo-fármaco não funciona assim. Os beta-receptores são alvos terapêuticos; isso não significa que qualquer estimulante seja um tratamento. Os receptores opioides são alvos terapêuticos; isso não significa que todos os agonismos opioides sejam benéficos. O mesmo princípio aplica-se aqui.
Em segundo lugar, os estudos mecanísticos não eliminam a preocupação epidemiológica. A investigação sobre enviesamento do recetor e desenho de ligandos coexiste, e não se opõe, às evidências de que a exposição ao cannabis pode agravar o risco de psicose em algumas pessoas, especialmente com produtos com elevado teor de THC, início precoce ou uso intenso. Um mecanismo do recetor pode explicar porque é que alguns compostos podem ser concebidos para melhores resultados. Não justifica afirmações públicas amplas de que “a ativação do CB1 é boa para a esquizofrenia”.
Em terceiro lugar, os leitores não devem tratar endocannabinoid, fitocannabinoid e ligandos sintéticos como intercambiáveis. A anandamida não é THC. Um modulador do CB1 com restrição periférica não é cannabis fumada. Um composto experimental de baixa eficácia e com viés de via não é um produto de consumo com elevada exposição. Estas categorias importam porque o comportamento do recetor depende da classe do ligando, da dose, da via de administração, do tempo e da exposição tecidular.
Por fim, a existência de medicamentos relacionados com cannabis aprovados não resolve as questões psiquiátricas. Até 2025, a FDA observa que um produto farmacológico derivado de cannabis e três produtos farmacológicos relacionados com cannabis foram aprovados. Esse facto mostra que a farmacologia dos cannabinoid pode tornar-se medicina sob condições definidas. Não significa que a estimulação não refinada do recetor seja segura, eficaz ou apropriada para a esquizofrenia.
A conclusão correta é mais rigorosa e menos reconfortante. A biologia do CB1 é relevante para a psicose porque o recetor está profundamente integrado em circuitos cerebrais que moldam saliência, cognição e inibição. Essa relevância apoia um desenho cuidadoso de fármacos, não extrapolações casuais. Na investigação da esquizofrenia, a farmacologia dos recetores é, ao mesmo tempo, um mapa de possibilidades e de riscos.
O que a ciência dos recetores significa para futuros medicamentos cannabinoid
A medicina cannabinoid está a afastar-se da velha ideia de que um fármaco útil precisa apenas de “atingir o CB1” ou “atingir o CB2”. Essa abreviação sempre foi incompleta. Agora é ativamente enganadora. CB1 e CB2 não são simples interruptores de ligar/desligar separados de forma nítida entre cérebro e sistema imunitário. São recetores com distribuições sobrepostas, mas irregulares, entre tipos celulares, conformações dinâmicas, preferências de acoplamento distintas e papéis variáveis ao longo dos estados de doença. Isto importa porque o mesmo ligando pode parecer terapêutico num tecido, intoxicante noutro e ineficaz num ensaio clínico se a população doente for demasiado ampla ou se o esquema posológico induzir dessensibilização.
Os riscos para a saúde pública não são menores. A Organização Mundial da Saúde estimou que 200 milhões de pessoas usaram cannabis em 2019, o equivalente a 4% da população mundial entre os 15 e os 64 anos. No entanto, a FDA dos EUA continua a referir que, até 2025, apenas um medicamento derivado de cannabis e três medicamentos relacionados com cannabis tinham sido aprovados. Essa diferença entre exposição generalizada e terapêuticas aprovadas limitadas é uma das razões pelas quais a farmacologia dos recetores é tão importante. Outra é a carga da doença: a epilepsia afeta cerca de 50 milhões de pessoas em todo o mundo, a esquizofrenia cerca de 24 milhões, e a dor crónica afeta quase 1 em cada 5 adultos nos Estados Unidos. Se a biologia dos cannabinoid for produzir medicamentos melhores, isso não acontecerá através de uma ativação grosseira dos recetores.
O trabalho fundador de Raphael Mechoulam, Lumír Hanuš e Allyn Howlett estabeleceu o campo moderno ao identificar a sinalização endocannabinoid e a farmacologia dos recetores. A fase seguinte é diferente. Trata-se menos de provar que CB1 e CB2 existem e mais de aprender a orientá-los com precisão.
| Conceito de desenho | O que procura melhorar | Porque o artigo diz que isso é importante |
|---|---|---|
| Seletividade de subtipo | Preferir CB1 ou CB2 de forma mais clara | O subtipo, por si só, não garante eficácia nem segurança |
| Ajuste de eficácia | Evitar ativação demasiado forte do recetor | Diferentes estados ativos podem alterar benefício, tolerância e efeitos adversos |
| Sinalização enviesada | Favorecer uma via intracelular em detrimento de outra | Pode ajudar a separar efeitos úteis de limitações |
| Modulação alostérica | Ajustar a resposta do recetor aos ligandos endógenos | Poderia preservar melhor a sinalização espacial e temporal normal |
| Restrição tecidular | Limitar a exposição a órgãos selecionados ou evitar a entrada no cérebro | Útil quando predominam os efeitos adversos centrais |
Seletividade, eficácia, modulação alostérica e viés
A lição mais importante da ciência recente dos recetores é que “ligação” não é o mesmo que “efeito”. Um ligando pode preferir CB1 em relação a CB2, ou vice-versa, mas isso é apenas o início. Também pode diferir na eficácia, isto é, na forma como estabiliza com maior ou menor intensidade os estados ativos do recetor. Pode favorecer a sinalização Gi/o em detrimento do recrutamento de beta-arrestina, ou promover mais a internalização do recetor do que uma sinalização sustentada à superfície celular. Essas diferenças podem alterar os resultados de forma drástica.
A revisão de 2026 da Frontiers in Chemical Biology sobre a estrutura dos recetores de cannabinoid torna este ponto claro: a seletividade de subtipo entre CB1 e CB2 depende de diferenças estruturais ao nível do recetor, que moldam a ligação do ligando, a eficácia da sinalização e a regulação do recetor. É por isso que os cannabinoid quimicamente semelhantes não se comportam de forma idêntica. Um phytocannabinoid, um endocannabinoid como a anandamida e um ligando sintético podem contactar a mesma família de recetores, produzindo, contudo, perfis downstream diferentes. O desenho de fármacos que ignora a eficácia e o viés da sinalização ficou agora atrás da evidência.
CB1 é o caso mais claro. É frequentemente discutido como o recetor responsável pelos efeitos psicoativos, o que é verdade, mas incompleto. CB1 também regula a transmissão sináptica, o processamento da dor, o apetite, os circuitos do stress e a sinalização cortical. O artigo de 2025 do American Journal of Psychiatry defende que a sinalização enviesada de CB1 é uma estratégia terapêutica plausível para a esquizofrenia. Esta é uma mudança marcante no campo. A esquizofrenia afeta aproximadamente 24 milhões de pessoas em todo o mundo, e o artigo não trata CB1 como um recetor a evitar a todo o custo. Em vez disso, trata CB1 como um alvo cujas consequências patológicas e terapêuticas podem depender das vias intracelulares que um ligando favorece. Isto é farmacologia clássica de GPCR, agora aplicada seriamente a terapêuticas baseadas em cannabinoid.
CB2 passou por uma reavaliação semelhante. A revisão de 2026 da Frontiers in Behavioral Neuroscience descreve “uma atualização dos últimos 3 anos”, mostrando um interesse crescente na sinalização de CB2 em perturbações do sistema nervoso central, especialmente quando estão envolvidas mecanismos neuroinflamatórios e neurodegenerativos. Isto não significa que CB2 seja um alvo anti-inflamatório mágico. Significa que a velha afirmação de que CB2 é irrelevante para o cérebro já não se ajusta à literatura. A expressão pode ser baixa em condições basais e aumentar em populações celulares específicas ou em contextos de doença. Um medicamento dirigido a CB2 pode, portanto, depender fortemente do momento, da patologia e do estado tecidual.
A modulação alostérica pode tornar-se uma das formas mais úteis de explorar estas distinções. Os agonistas ortostéricos frequentemente induzem uma ativação ampla do recetor, o que aumenta o risco de efeitos adversos, tolerância e fraca seletividade contextual. Em contraste, os moduladores alostéricos podem ajustar a forma como os recetores respondem aos ligandos endógenos, em vez de forçarem uma ativação máxima. Em princípio, isso poderia preservar os aspetos espaciais e temporais da sinalização endocannabinoid normal. Trata-se de uma estratégia apelativa para CB1, onde a agonização direta frequentemente esbarra em efeitos adversos centrais, e para CB2, onde a sobrerregulação associada à doença pode tornar a modulação dependente do contexto mais atrativa do que a estimulação constante. Mas “apelativa” não significa “comprovada”. Muitas ideias alostéricas que parecem elegantes in vitro falham quando entram em jogo a reserva do recetor, a heterogeneidade tecidual e a farmacocinética.
Porque é que a biologia estrutural e em rede orienta agora o desenho de fármacos
O centro de gravidade do campo deslocou-se do simples mapeamento dos recetores para a análise dinâmica dos estados do recetor. É aqui que a biologia estrutural se torna importante. A revisão de 2026 da Frontiers e o recente estudo indexado em PubMed sobre o mecanismo dinâmico da seletividade de subtipo contestam o modelo de chave-fechadura. A seletividade não é apenas uma questão de um ligando encaixar melhor num bolso de um recetor. Os recetores “respiram”. Os ligandos estabilizam conjuntos de conformações. Os endocannabinoid podem alcançar preferência de subtipo através de interações dinâmicas que alteram a forma como CB1 ou CB2 se movem entre estados inativos, ativos e competentes para sinalização.
Essa mudança de pensamento tem consequências práticas. Um químico medicinal já não pergunta apenas: “Este composto liga-se a CB1 ou CB2?” As melhores perguntas são: Que conformações ele estabiliza? Promove mais Gi/o do que beta-arrestina? Desencadeia internalização rápida? Comporta-se de forma diferente em neurónios, microglia, hepatócitos ou nociceptores periféricos? Pode ser restringido à periferia? Coopera com o tónus do ligando endógeno que aumenta apenas durante inflamação ou lesão?
A biologia em rede estende ainda mais esta abordagem. Uma análise integrada recente indexada em PubMed identificou CB1 e CB2 como nós altamente influentes no sistema endocannabinoid e associou a sinalização dos recetores a vias metabólicas mais amplas. Isso é importante porque a farmacologia do endocannabinoid nunca se resume apenas aos recetores. A síntese, o transporte e a degradação da anandamida e do 2-AG moldam a exposição aos recetores. O mesmo acontece com o metabolismo lipídico, os mediadores inflamatórios e a comunicação cruzada com outros sistemas GPCR e canais iónicos. Um ligando que parece limpo num ensaio recombinante pode comportar-se de forma muito diferente em tecido vivo, onde FAAH, MAGL, vias das prostaglandinas e a densidade local dos recetores variam.
Esta perspetiva sistémica também ajuda a explicar porque é que a translação clínica tem sido irregular. Alguns insucessos terão resultado de compostos inadequados. Outros terão derivado de modelos de doença excessivamente simplificados. Se um ensaio recruta doentes cujos sintomas partilham um rótulo, mas não uma assinatura endocannabinoid, o resultado médio pode parecer dececionante, mesmo que um subgrupo biologicamente definido possa beneficiar. O sucesso futuro dependerá não só da química, mas também da estratificação: que doentes, que fase da doença, que tecidos, que estado do recetor, que tónus endógeno.
Expectativas realistas para a próxima geração de terapêuticas
A próxima geração de medicamentos cannabinoid provavelmente será mais estreita, mais seletiva e menos romântica do que sugere o debate público. Isso é uma boa notícia. Medicamentos melhores surgem muitas vezes quando um campo abandona explicações que servem para tudo.
Alguns progressos já são visíveis. A restrição periférica de CB1 continua a ser atrativa para indicações de dor ou metabólicas porque visa reduzir os efeitos adversos centrais. Compostos dirigidos a CB2 continuam a ser plausíveis para condições inflamatórias, neuropáticas e neuroimunes, especialmente se a expressão associada à doença criar janelas terapêuticas. Ligandos CB1 enviesados podem revelar utilidade em doenças neuropsiquiátricas, se conseguirem separar os efeitos de circuito desejados da intoxicação, ansiedade ou tolerância indesejadas. Os moduladores alostéricos podem oferecer um controlo mais subtil do que os agonistas diretos. As abordagens dirigidas às enzimas, que alteram indiretamente o tónus endocannabinoid, podem ainda ser valiosas, embora tragam os seus próprios riscos e não contornem a complexidade dos recetores.
Ainda assim, o realismo é essencial. Seletividade não garante sucesso. Um agonista de CB2 altamente seletivo pode falhar se a expressão de CB2 na doença-alvo for demasiado variável ou se a sinalização downstream não for, na realidade, o principal motor dos sintomas. O viés medido num ensaio pode não prever o viés em tecido humano. Os instantâneos estruturais são poderosos, mas continuam a ser instantâneos. A restrição tecidual pode reduzir algumas limitações e criar outras. E os phytocannabinoid não devem ser tratados como substitutos de toda a farmacologia dos cannabinoid; endocannabinoid, compostos de origem vegetal e ligandos sintéticos podem produzir comportamentos recetoriais muito diferentes dependendo da dose, do momento e do contexto de sinalização.
Assim, a previsão serena é esta: a futura medicina baseada em cannabinoid provavelmente melhorará por se tornar mais precisa, e não por se tornar mais amplamente ativadora. Os caminhos mais credíveis combinam seletividade de recetor, viés de sinalização, controlo alostérico, direcionamento tecidual e melhor estratificação de doentes. O que se sabe é substancial: CB1 e CB2 são centros dinâmicos de sinalização, o estado estrutural importa e o contexto da doença importa. O que permanece incerto é se essa clareza mecanística se traduzirá em ganhos clínicos reproduzíveis na dor, na doença psiquiátrica, na epilepsia, na neurodegeneração e na doença inflamatória. O campo continua a ser clinicamente importante porque a necessidade não satisfeita é grande, a biologia é real e o modelo simplista anterior deu finalmente lugar a uma farmacologia capaz de explicar tanto a promessa como a desilusão.
Referências
- [1]Cannabis (marijuana). WHO Fact Sheet, 2024. https://www.who.int/news-room/fact-sheets/detail/cannabis-(marijuana)
- [2]Epilepsy. WHO Fact Sheet, 2024. https://www.who.int/news-room/fact-sheets/detail/epilepsy
- [3]Schizophrenia. WHO Fact Sheet, 2022. https://www.who.int/news-room/fact-sheets/detail/schizophrenia
- [4]FDA Regulation of Cannabis and Cannabis-Derived Products, Including Cannabidiol (CBD). FDA Public Health Focus, 2025. https://www.fda.gov/news-events/public-health-focus/fda-regulation-cannabis-and-cannabis-derived-products-including-cannabidiol-cbd
- [5]The Health Effects of Cannabis and Cannabinoids: The Current State of Evidence and Recommendations for Research. National Academies Press, 2017. https://nap.nationalacademies.org/catalog/24625/the-health-effects-of-cannabis-and-cannabinoids-the-current-state